Adrian Buganza Tepole, PhD
Associate Professor of Mechanical Engineering, Biomedical Engineering (by courtesy), Purdue University
Introduction
Work on soft tissue constitutive modeling spans multiple decades. Much progress has been made but, of course, there are many challenges that remain. This journal club post is not intended to cover all the history of soft tissue constitutive modeling, for which there are excellent reviews and books, e.g. [1-3]. Rather, this post will focus on recent research directions in the literature, primarily over the last 5 years: i) modeling of dissipative mechanisms, i.e. viscoelasticity and damage; ii) coupled problems at the interface of biology-mechanics; iii) emerging topics in brain, lung, and reproductive biomechanics.
Constitutive modeling of tissues beyond hyperelasticity
The role of artificial intelligence and machine learning (ML) starting with hyperelasticity
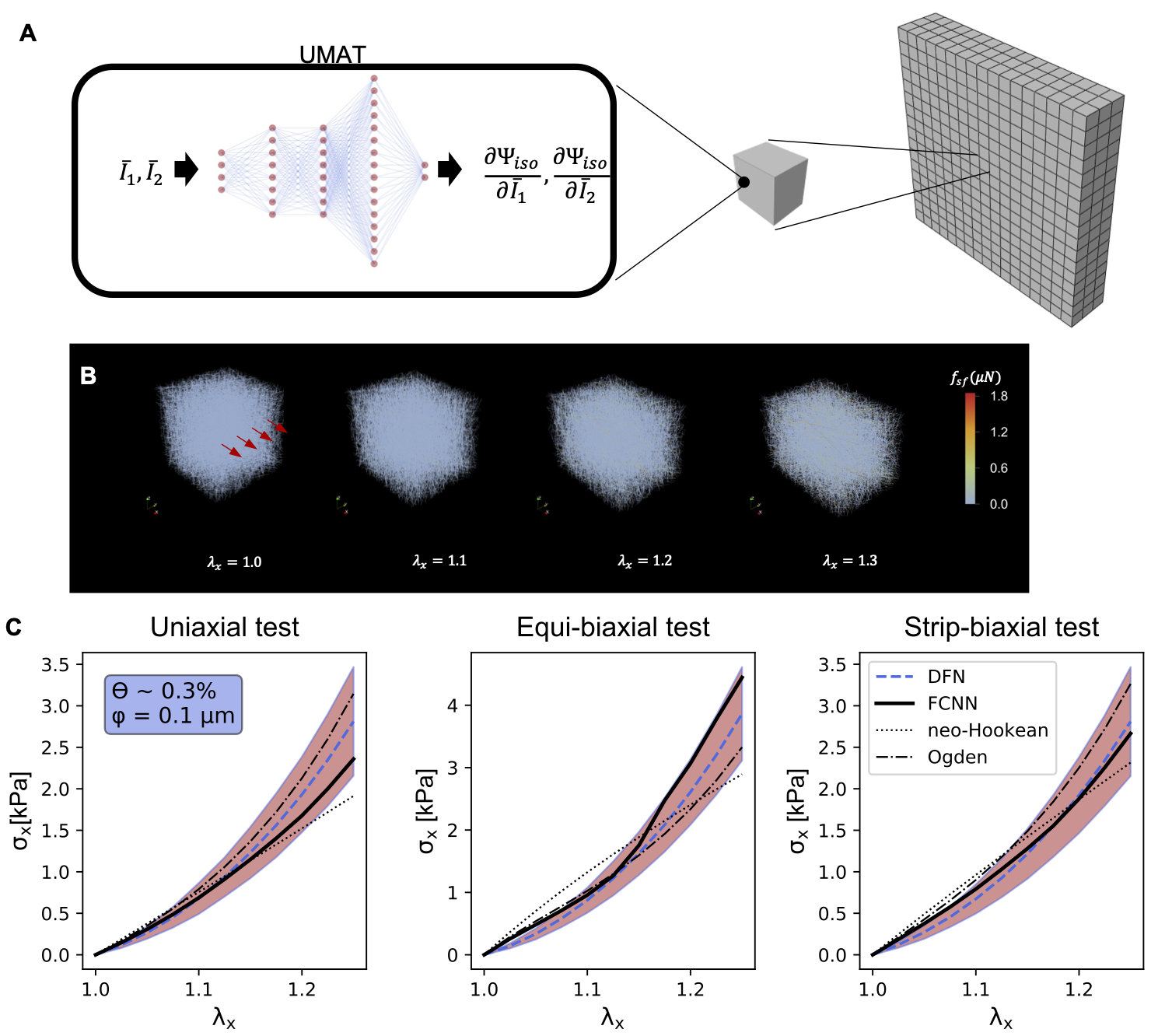
Hyperelastic modeling of soft tissue has been done since the pioneering work by Fung and collaborators seven decades ago [1]. Over the years, one avenue of research has been to include microstructure information as part of the strain energy function [3]. Even today, microstructure-based modeling of soft tissues continues to receive attention, especially for tissues that have only attracted robust research interest recently, e.g. in reproductive or brain biomechanics [4-7]. For both new tissues of interest as well as in more established fields such as cardiovascular biomechanics, ML has contributed to new types of modeling, starting within the context of hyperelasticity. Similar to the development of closed-form models by experts, ML methods can be based on stress-strain data [8, 9] only, or microstructure-based [10]. In contrast to micromechanics derivations (collagen recruitment functions, micro-sphere formulations), ML tools enable direct homogenization of detailed simulations of representative volume elements (RVE) [11, 12]. A thorough discussion of ML tools for constitutive modeling are out of the scope of this blog, but the interested reader is referred to the excellent review by Jin et al [13].
Fig 1. ML can be used to directly homogenize the response from detailed microstructure models (A). For example, we have developed different fully connected neural network approaches to learn the response of discrete fiber network models (B), under arbitrary deformations and across microstructures defined in terms of fiber diameters and volume fractions. The homogenized response predicted with the ML method outperforms analytical models and works over a range of microstructures (C) [11].
New theories for viscoelasticity based on fractional calculus and kinetics of reactive mixtures
Dissipative mechanisms in biological materials have also been modeled before. However, there are several recent efforts along this avenue of investigation that we want to highlight. The quasi-linear viscoelastic (QLV) formulation by Fung has remained extremely popular over the years [1]. Surprisingly, the thermodynamic consistency of this approach has only been proven recently [14]. In its original form, the QLV was difficult to implement into finite element formulations because of the memory cost of storing the entire stress history in order to compute the convolution integral with the relaxation function over all time. However, restricting the relaxation function to a sum of exponential decays enabled an efficient finite element implementation (only one internal variable per time step per exponential needs to be stored) [15]. The downside of dealing with the sum of exponentials is the limited type of viscoelastic behavior that can be captured.
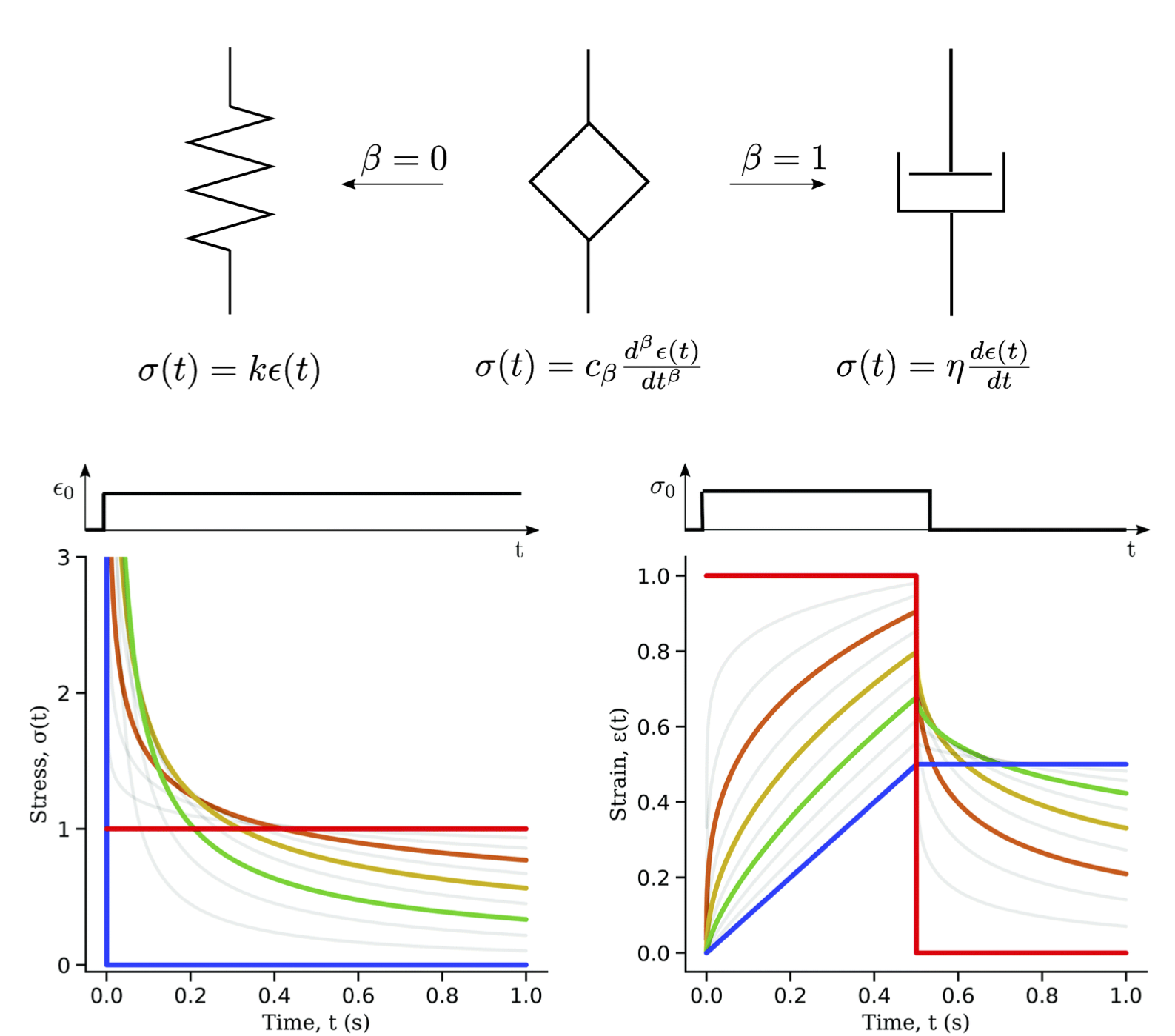
Fractional calculus has emerged as a new framework for viscoelasticity of tissue which is not limited to exponential decay [16]. A good overview of fractional viscoelasticity for one-dimensional examples that’s a good entry point in this topic is [17]. In the one-dimensional case it becomes clear that the fractional order derivative of the springpot element interpolates smoothly between the behavior of a spring and that of a dashpot from the classical rheological models. Another new perspective for dissipation of energy in tissues is reactive viscoelasticity [18]. This theory has origins in similar observations made for rubbers [19]. In reactive viscoelasticity, reaction kinetics take the protagonist role. Continuous formation and breaking of bonds (fiber-fiber or intra-fiber crosslinks and other strong and weak interactions), occurring at different deformations of the material, can dissipate energy. They key idea is that bond formation occurs at the current state of stress of the body; a new bond does not carry stress. The reactive mixture model, much like fractional calculus approach, can naturally capture non-exponential relaxation which is typical of soft tissue.
Fig 2. Fractional derivatives can be used to smoothly transition between a spring and classical dashpot used in rheological models of viscoelasticity. The bottom plots show response under stress relaxation or creep. The red curve is the spring limit and the blue plot is the dashpot limit. The figure is taken with permission from [17].
ML enhances existing viscoelasticity frameworks
Similar to hyperelastic modeling, ML has also contributed to more flexible and accurate constitutive models of viscoelasticity. The most straightforward use of ML is automatic systems identification of models and parameters. For instance, Wang and collaborators learned QLV models of muscle with recurrent neural networks [20]. For non-equilibrium processes at large deformations, a natural approach is to construct energy and dissipation potentials with neural networks. We have used neural ordinary differential equations to build convex potentials that automatically satisfy polyconvexity and the second law of thermodynamics [21]. ML methods extending closed-form models of large deformation, anisotropic viscoelasticity have increased flexibility and higher accuracy [22, 23].
Understanding damage and fatigue failure in soft tissue needs new multiscale theories that complement new experimental observations
Energy dissipation due to damage has also received some attention in the past, not just in the last five years, but there are good developments in recent years that are worth discussing. A good review of the state of the field by 2016 is that by Li [24]. Constitutive models of soft tissue damage build upon a very extensive theory of damage for large deformation elasticity because of the focus in rubber elasticity in the second half of the twentieth century. The so-called Mullins effect entails the reduction of the strain energy capacity as a function of the previously experience maximum deformation [25]. Modeling rubber damage was advanced, among others, by Simo and Ogden [26-28]. These isotropic models, termed pseudo-elasticity and continuum damage mechanics, have also been used in the context of tissue mechanics. Even though the isotropic models were able to capture the reduction in strain energy with respect to damage accumulation, they were not able to reproduce the permanent set phenomenon which is also seen in tissues. Microstructure-based approaches for rubber captured anisotropic damage and explained permanent set as a result of this induced anisotropy even if the initial material was isotropic [29]. In the last five years, microstructure-based models have been developed to model damage in biological materials [30-32]. The microscale models typically consider distributions of fiber orientation and fiber recruitment, model individual fiber as a relatively simple material (such as neo-Hookean), and use a relatively simple damage criteria for individual fibers (strain-dependent failure, fiber sliding). While effective, these damage formulations tend to be local, driven by internal variables at integration points of a finite element implementation, and can thus result in localization and mesh dependence. Non-local damage formulations are now being developed for soft tissues to deal with these problems [33, 34].
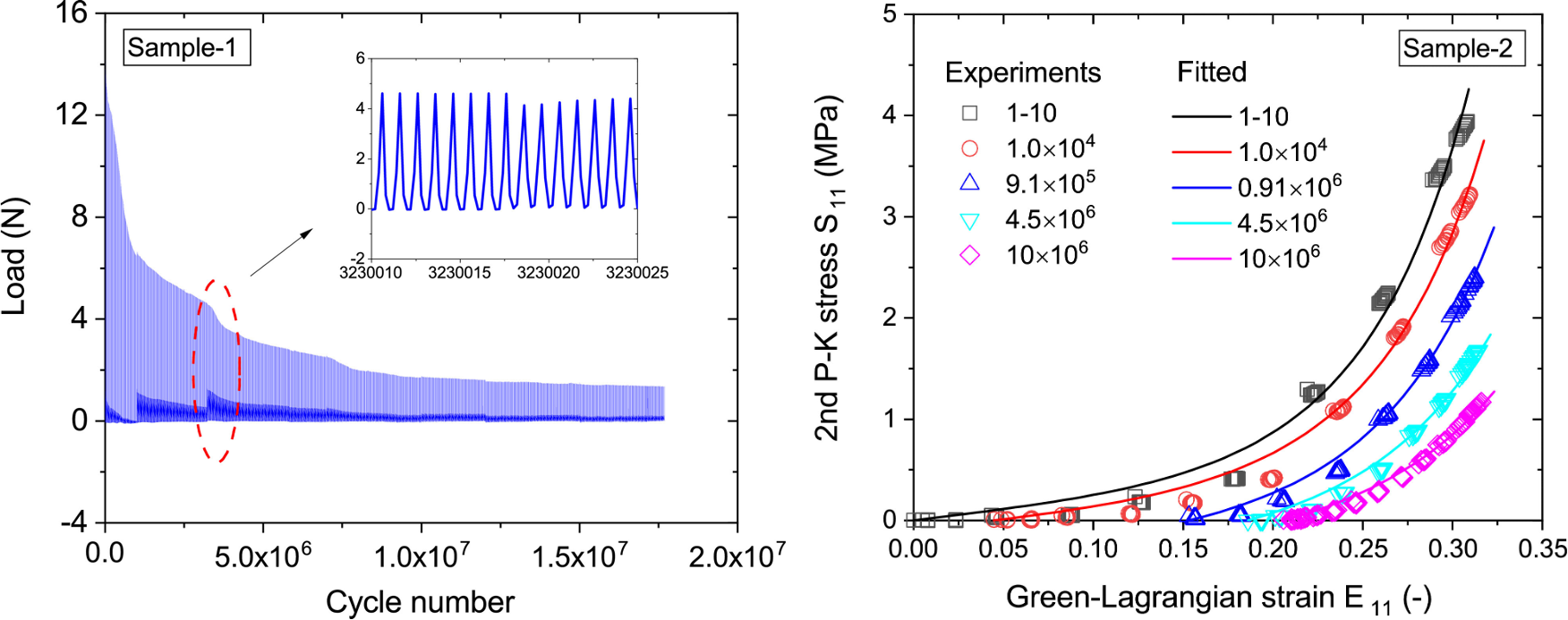
Modeling acute tissue damage is important for applications such as subcutaneous drug delivery and surgery, but damage can accrue over longer time scales. This is the case for tissues that undergo continuous cyclic loading, for instance engineering tissue or for overuse injury in musculoskeletal tissues [35, 36]. Remarkably, many tissues operate for millions of cycles without mechanical injury because of the ability of living tissue to repair itself. Tissue engineered heart valve replacements and other tissue engineered constructs lack fatigue resistance [37]. In line with the damage models discussed above, fatigue failure of collagenous tissues has been modeled based on microstructure arguments. Operating directly at the continuum level, Zhang and Sacks modeled fatigue in a microstructure-based approach where collagen recruitment functions were updated as a function of deformation to capture permanent set [38]. In their model, the evolution of the recruitment functions had a kinematic coupling (fibers tended to deform permanently as a function of experienced strain) but with a reaction rate akin to the reactive viscoelasticity models. Martin and Sun also follow a microstructure-motivated strategy, but adopted the damage mechanics framework by Simo to model progressive damage of individual fibers as a function of maximum experienced equivalent strain [39]. Interestingly, as more detailed models involving more than one phenomenon are deployed, emergent features can be observed. Several recent studies have shown that coupling of damage mechanics with other forms of energy dissipation are needed to explain experimentally observed fatigue failure. For instance, Henderson et al. show that damage models coupled with viscous behavior using the QLV formulation are needed to explain observations [40]. Zitnay and co-workers also show coupling between damage and other energy dissipation mechanisms, in their case poroelasticity [41].
Fig 3. Typical damage accumulation in soft tissue. Subjected to displacement-driven deformation, load will decrease over millions of cycles of loading. The resulting stress-strain response shows the damage degradation for samples tested after different amount of fatigue. Taken with permission from [37].
Fatigue failure has gained increasing attention in other fields besides tissue modeling, most notably for hydrogels [42]. Therefore, an important task for researchers in biomechanics is to become familiar with developments in hydrogel fatigue and vice versa, in order to continue moving this avenue of research forward.
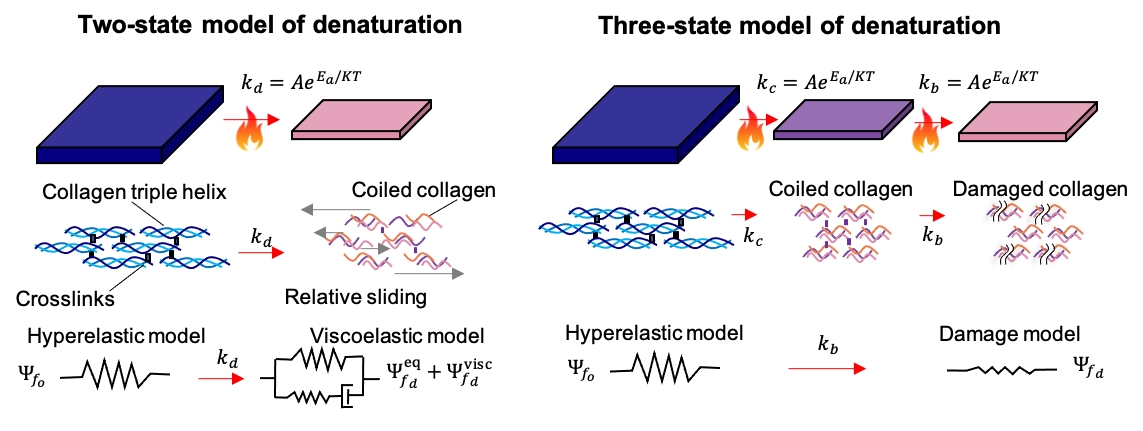
Despite of the success modeling damage and fatigue failure of tissues at the macroscopic scale, one of the limiting factors for new constitutive model development is the lack of experiments that can point to the precise microscopic events leading to the observed macroscale response. For instance, we proposed different mechanisms to explain the thermomechanics of skin building on the microstructure arguments that have been reviewed here [38, 39, 43, 44]. New imaging techniques and manipulation of small tissue samples are revealing the multiscale couplings behind tissue failure [41]. To match the experimental advances, similar multiscale models are needed [45]. For example, Dhume et al. are working to scale up failure in discrete fiber network models to continuum level descriptions [46]. This type of multiscale integration seems a perfect opportunity for ML methods, just as ML has been used to homogenize hyperelastic response out of RVE simulations. For damage, fatigue, and fracture, coupling from the molecular scale, to the fiber network, to the tissue level is needed.
Fig 4. Damage due to heat can be modeled with the different microstructure-based models reviewed here [38, 39, 43, 44]. Namely, reaction rates for crosslinks and other, weaker, inter- and intra- collagen fiber interactions, coupled to damage models or viscoelasticity models based on fiber sliding, and stress-free lengths of intact and denatured collagen can combine to capture thermomechanics damage. However, new experiments and detailed multiscale simulations from the molecular scale up to the tissue are needed [41, 45].
Tight integration of continuum mechanics and computational biology
Higher order theories of growth and remodeling
Thinking about the mechanics of tissue immediately brings to mind the ageing, remodeling, and healing that is associated with living systems. This aspect of tissue mechanics has also received attention for several decades. Rodriguez et al. were some of the first to propose a continuum mechanics framework for growth (addition of mass), in response to mechanical loading [47]. The multiplicative split of the deformation gradient into growth and elastic contributions, inspired by large deformation plasticity, has continued to be used to describe a variety of tissues over the years, including recent examples such as skin and brain growth/atrophy [48, 49]. Humphrey and Rajagopal proposed a theory based on constrained mixtures [50] that has also continued to be used in many practical applications up to today [51].
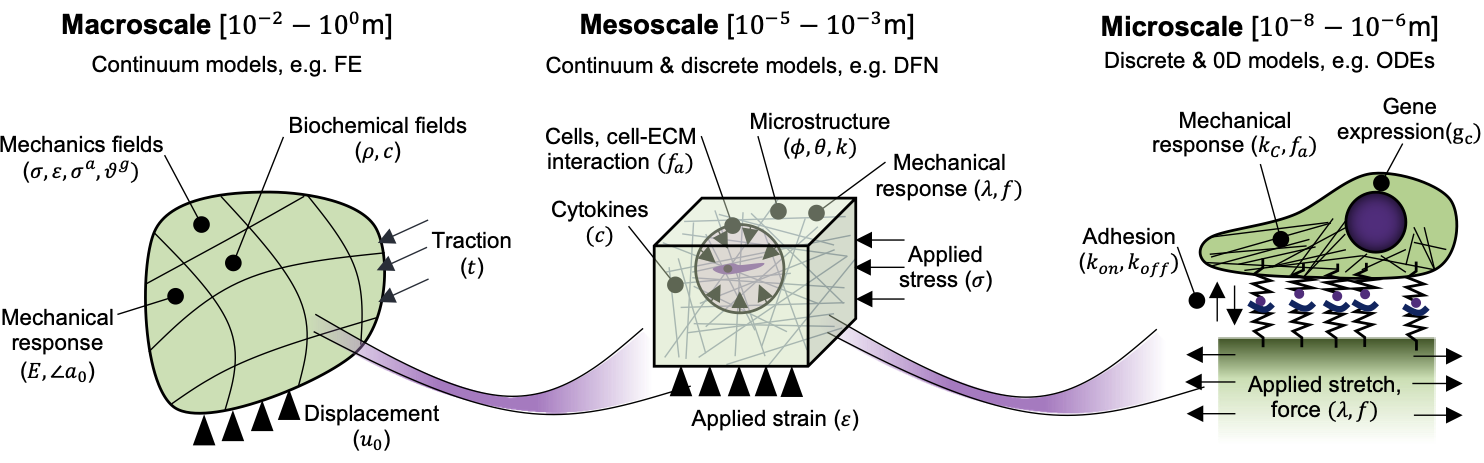
What are the novel developments around these theories in recent years, beyond their application to predict disease or treatment in a particular organ system? Ignoring the biological aspects for a moment, some innovations in growth and remodeling theories are consideration of higher order derivatives and non-local effects. For example, related to the discussion of gradient-enhanced damage models, similar frameworks can be used to model the reverse process: healing of soft tissue [52, 53]. Many self-healing polymer mechanics theories have been developed and could be adapted to the open-system thermodynamics of living tissue. Zuo and co-authors proposed a healing model based on thermodynamical considerations along this line of thought [54]. In crystal plasticity, the multiplicative split of the deformation gradient has been accompanied by a thorough analysis of the geometry of defects of the crystal lattice. What is the equivalent geometry for growing tissue? We were interested in using metrics of incompatibility to describe the residual stresses arising from growth [55]. However, multiscale descriptions linking changes in microstructure up to the continuum level are needed to properly understand and model a defect theory of growth. Work by Gacek et al [56] is bridging discrete fiber network mechanics to the tissue scale formulation of growth in terms of the multiplicative split of the deformation gradient. Here, once again, there is room for ML tools to link detailed models of cells interacting with their immediate environment, up to the continuum level models.
Fig 5. Modeling of tissue growth and remodeling requires integration across spatial scales, from the cell adhesion modeling and corresponding gene expression, to the mesoscale fiber networks, to the tissue level mechanics [57].
Computational systems mechanobiology
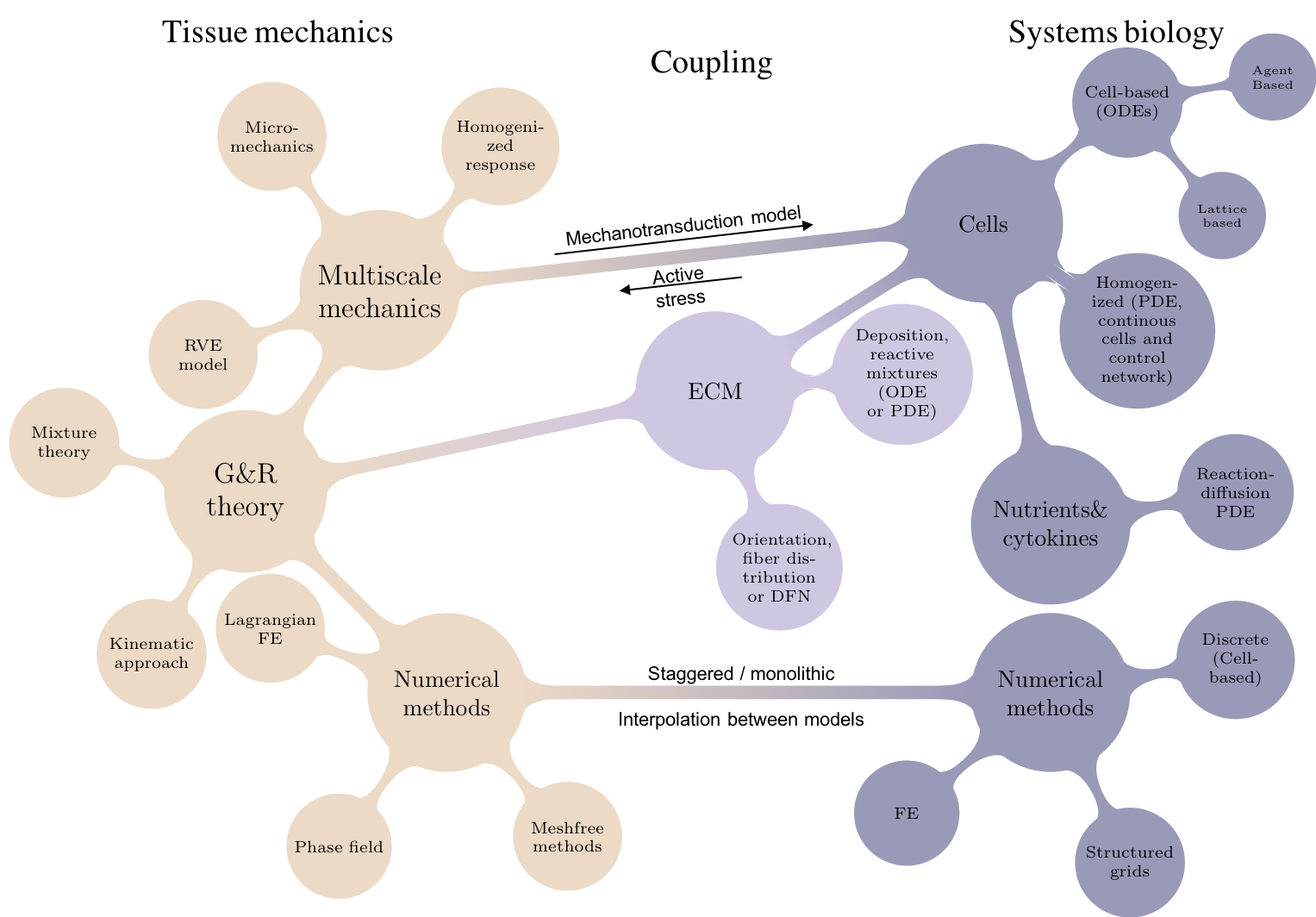
Another major trend is the integration of continuum mechanics to more detailed biological systems modeling [58]. Growth and remodeling theories based almost exclusively on mechanics variables can successfully predict the quantities of interest, e.g. skin growth in tissue expansion [48]. Yet, in some cases this is not enough due to the intricacies of cell-regulatory networks or systemic changes in physiology that have downstream effects in tissue mechanics. Pregnancy is a primary example [59]. Hormonal changes during pregnancy lead to heart remodeling. Understanding these effects requires explicit modeling of the intracellular signaling that transforms the chemical cues to the cellular activity leading to remodeling. The field of systems biology has produced models, often based on either logic decision trees or systems of ordinary differential equation (ODE) or stochastic differential equations (SDE), that can capture concentration of chemical signals or normalized gene and protein expression dynamics [60]. For soft tissue, dynamic models of cells such as fibroblasts or cardiomyocytes are needed, including variables for genes or proteins that can be linked to remodeling such as cell proliferation and collagen deposition [61].
Examples of comprehensive models linking biology and mechanics abound for cardiovascular tissues in the literature of the past five years. Irons and co-authors, for instance, have integrated a dynamic model of fibroblast and smooth muscle cell mechanobiology [62] to a continuum mechanics model or artery remodeling [63]. Other examples from the last couple of years along similar lines are [64-66]. Other soft tissues for which this integrative approach is being applied are cartilage [67], and skin [68].
There are some interesting fundamental questions arising as a result of coupling continuum mechanics of growth and remodeling with dynamical systems, regardless of the specific intricacies of the biology of different tissues. For instance, dynamical systems are characterized by the beauty of instabilities, multi-stable states, and oscillations. Remarkably, in biology, there are particular patterns or motifs that appear over and over [69, 70]. What are the emergent mechanics when these motifs are coupled to, for instance, permanent deformation (growth)?
Opportunities for ML models of growth and remodeling
One last point to conclude this section, similar to the preceding ones, is regarding ML. How could we ignore it? Even though for the systems biology side of things there have been indeed multiple efforts to integrate ML to improve constitutive models of cell signaling [71], there has been relatively little on the side of growth and remodeling biomechanics. One interesting example is by Liu et al. who use reinforcement learning to discover growth constitutive equations [72].
Fig 6. Graphical overview of the variables of interest, theories, and numerical methods needed to fully integrate the observed mechanics of growth and remodeling to computational systems biology [58].
New tissues come into focus
Brain, lung, and reproductive biomechanics
Undeniably, musculoskeletal and cardiovascular tissues have had a lot of well-deserved attention in the field of tissue mechanics. Other tissues have made sporadic appearances in the literature, e.g. eyes and skin (my favorite). But few could argue that, after a global pandemic of a respiratory disease, lung biomechanics has come to the front. Without extending this review too much more, we would like to point out the reader to new developments in this area, specifically regarding constitutive model of lung tissue e.g.: [73-75]. A common thread joining these efforts is the focus on RVE models that can be scaled up to obtain tissue level parenchyma behavior. Due to the unique microscale geometry of the parenchyma, there are emergent relationships for the multiscale mechanics that had not been observed in the multiscale modeling of other tissues. This is not entirely surprising, as every tissue has a unique mechanical function tuned over millions of years of evolution. Thus, even as we try paradigms that have been put forward for other tissues, their application to new tissues requires modeling innovation and results in new mechanics knowledge. The brain is also in everybody’s head. With an ageing global population, brain atrophy and dementia appear routinely in news headlines, e.g. whether the latest Alzheimer’s drugs clear FDA approval or not. Constitutive model of brain is, understandably, a very active field of research today. Once again, without extending the review much more, we would just like to point out a couple of excellent efforts in this area: [5, 49, 76]. In line with the trends we have already covered, constitutive modeling of brain in the last five years has been focused on incorporating microstructure knowledge, incorporating dissipative mechanisms such as viscoelasticity, and understand brain atrophy as a coupling between systems biology and the mechanics of growth and remodeling. Lastly, we would like to give a shout out to the field of reproductive biomechanics. It was only in 2015 that the NIH released the SABV policy which made mandatory the inclusion of sex as a biological variable. Clearly, the history of science is not free of bias. It is not surprising, then, that constitutive modeling of the female reproductive system has not received enough attention. Thankfully, it is never late to address the gaps, and researchers in this field have been very active in the last five years. A couple of examples of constitutive modeling of cervix and uterus tissues are: [7, 77, 78].
Discussions
This review has highlighted some trends in the last five years of soft tissue constitutive model research.
- What did we miss that should be top of the list when thinking about recent advances in constitutive models of soft tissue?
- In addition to the three specific tissues highlighted here, what is your favorite tissue and can you point to recent papers that focus on that tissue?
- The integration across scales and between systems biology and mechanics was deemed important to move the field forward, potentially aided by ML. What are the challenges you foresee for building more and more complete models of soft tissue? Both experimental as well as modeling challenges.
- What should be investigated to advance soft tissue constitutive modeling that is not currently receiving enough attention? Do you have predictions for emerging topics of interest that are not yet the center of attention in soft tissue modeling?
References
1. Fung, Y.-c., Biomechanics: mechanical properties of living tissues. 2013: Springer Science & Business Media.
2. Humphrey, J.D. and S.L. Delange, An introduction to biomechanics. Solids and Fluids, Analysis and Design. Springer, Heidelberg, 2004.
3. Kassab, G.S. and M.S. Sacks, Structure-based mechanics of tissues and organs. 2016: Springer.
4. Cosentino, F., S. Sherifova, G. Sommer, G. Raffa, M. Pilato, S. Pasta, and G.A. Holzapfel, Regional Biomechanical Characterization of Human Ascending Aortic Aneurysms: Microstructure and Biaxial Mechanical Response. Acta Biomaterialia, 2023.
5. Budday, S., M. Sarem, L. Starck, G. Sommer, J. Pfefferle, N. Phunchago, E. Kuhl, F. Paulsen, P. Steinmann, and V. Shastri, Towards microstructure-informed material models for human brain tissue. Acta Biomaterialia, 2020. 104: p. 53-65.
6. Nikpasand, M., R.R. Mahutga, L.M. Bersie-Larson, E. Gacek, and V.H. Barocas, A hybrid microstructural-continuum multiscale approach for modeling hyperelastic fibrous soft tissue. Journal of Elasticity, 2021. 145(1-2): p. 295-319.
7. Shi, L., L. Hu, N. Lee, S. Fang, and K. Myers, Three-dimensional anisotropic hyperelastic constitutive model describing the mechanical response of human and mouse cervix. Acta Biomaterialia, 2022. 150: p. 277-294.
8. Tac, V., V.D. Sree, M.K. Rausch, and A.B. Tepole, Data-driven modeling of the mechanical behavior of anisotropic soft biological tissue. Engineering with Computers, 2022. 38(5): p. 4167-4182.
9. Linka, K. and E. Kuhl, A new family of Constitutive Artificial Neural Networks towards automated model discovery. Computer Methods in Applied Mechanics and Engineering, 2023. 403: p. 115731.
10. Holzapfel, G.A., K. Linka, S. Sherifova, and C.J. Cyron, Predictive constitutive modelling of arteries by deep learning. Journal of the Royal Society Interface, 2021. 18(182): p. 20210411.
11. Leng, Y., V. Tac, S. Calve, and A.B. Tepole, Predicting the mechanical properties of biopolymer gels using neural networks trained on discrete fiber network data. Computer Methods in Applied Mechanics and Engineering, 2021. 387: p. 114160.
12. Kakaletsis, S., E. Lejeune, and M.K. Rausch, Can machine learning accelerate soft material parameter identification from complex mechanical test data? Biomechanics and Modeling in Mechanobiology, 2023. 22(1): p. 57-70.
13. Jin, H., E. Zhang, and H.D. Espinosa, Recent Advances and Applications of Machine Learning in Experimental Solid Mechanics: A Review. arXiv preprint arXiv:2303.07647, 2023.
14. Berjamin, H., M. Destrade, and W.J. Parnell, On the thermodynamic consistency of quasi-linear viscoelastic models for soft solids. Mechanics Research Communications, 2021. 111: p. 103648.
15. Puso, M. and J. Weiss, Finite element implementation of anisotropic quasi-linear viscoelasticity using a discrete spectrum approximation. 1998.
16. Zhang, W., A. Capilnasiu, G. Sommer, G.A. Holzapfel, and D.A. Nordsletten, An efficient and accurate method for modeling nonlinear fractional viscoelastic biomaterials. Computer methods in applied mechanics and engineering, 2020. 362: p. 112834.
17. Bonfanti, A., J.L. Kaplan, G. Charras, and A. Kabla, Fractional viscoelastic models for power-law materials. Soft Matter, 2020. 16(26): p. 6002-6020.
18. Ateshian, G.A., C.A. Petersen, S.A. Maas, and J.A. Weiss, A Numerical Scheme for Anisotropic Reactive Nonlinear Viscoelasticity. Journal of Biomechanical Engineering, 2023. 145(1): p. 011004.
19. Green, M.S. and A.V. Tobolsky, A new approach to the theory of relaxing polymeric media. The Journal of chemical physics, 1946. 14(2): p. 80-92.
20. Wang, L.M., K. Linka, and E. Kuhl, Automated model discovery for muscle using constitutive recurrent neural networks. Journal of the Mechanical Behavior of Biomedical Materials, 2023. 145: p. 106021.
21. Taç, V., M.K. Rausch, F.S. Costabal, and A.B. Tepole, Data-driven anisotropic finite viscoelasticity using neural ordinary differential equations. Computer Methods in Applied Mechanics and Engineering, 2023. 411: p. 116046.
22. Reese, S. and S. Govindjee, A theory of finite viscoelasticity and numerical aspects. International journal of solids and structures, 1998. 35(26-27): p. 3455-3482.
23. Nguyen, T., R. Jones, and B. Boyce, Modeling the anisotropic finite-deformation viscoelastic behavior of soft fiber-reinforced composites. International Journal of Solids and Structures, 2007. 44(25-26): p. 8366-8389.
24. Li, W., Damage models for soft tissues: a survey. Journal of Medical and Biological Engineering, 2016. 36: p. 285-307.
25. Mullins, L., Effect of stretching on the properties of rubber. Rubber chemistry and technology, 1948. 21(2): p. 281-300.
26. Govindjee, S. and J. Simo, A micro-mechanically based continuum damage model for carbon black-filled rubbers incorporating Mullins' effect. Journal of the Mechanics and Physics of Solids, 1991. 39(1): p. 87-112.
27. Simo, J. and J. Ju, Strain-and stress-based continuum damage models—I. Formulation. Mathematical and Computer Modelling, 1989. 12(3): p. 378.
28. Ogden, R.W. and D.G. Roxburgh, A pseudo–elastic model for the Mullins effect in filled rubber. Proceedings of the Royal Society of London. Series A: Mathematical, Physical and Engineering Sciences, 1999. 455(1988): p. 2861-2877.
29. Göktepe, S. and C. Miehe, A micro–macro approach to rubber-like materials. Part III: The micro-sphere model of anisotropic Mullins-type damage. Journal of the Mechanics and Physics of Solids, 2005. 53(10): p. 2259-2283.
30. Rausch, M.K. and J.D. Humphrey, A microstructurally inspired damage model for early venous thrombus. Journal of the mechanical behavior of biomedical materials, 2016. 55: p. 12-20.
31. Toaquiza Tubon, J.D., O. Moreno-Flores, V.D. Sree, and A.B. Tepole, Anisotropic damage model for collagenous tissues and its application to model fracture and needle insertion mechanics. Biomechanics and Modeling in Mechanobiology, 2022. 21(6): p. 1-16.
32. Li, K. and G.A. Holzapfel, Multiscale modeling of fiber recruitment and damage with a discrete fiber dispersion method. Journal of the Mechanics and Physics of Solids, 2019. 126: p. 226-244.
33. Zuo, D., S. Avril, C. Ran, H. Yang, S.J. Mousavi, K. Hackl, and Y. He, Sensitivity analysis of non‐local damage in soft biological tissues. International Journal for Numerical Methods in Biomedical Engineering, 2021. 37(3): p. e3427.
34. Rausch, M.K., G.P. Sugerman, S. Kakaletsis, and B. Dortdivanlioglu, Hyper-viscoelastic damage modeling of whole blood clot under large deformation. Biomechanics and Modeling in Mechanobiology, 2021. 20(5): p. 1645-1657.
35. Linka, K., M. Hillgärtner, and M. Itskov, Fatigue of soft fibrous tissues: Multi-scale mechanics and constitutive modeling. Acta biomaterialia, 2018. 71: p. 398-410.
36. Pouca, M.V., P. Areias, S. Göktepe, J. Ashton-Miller, R.N. Jorge, and M. Parente, Modeling permanent deformation during low-cycle fatigue: Application to the pelvic floor muscles during labor. Journal of the Mechanics and Physics of Solids, 2022. 164: p. 104908.
37. Dong, H., M. Liu, C. Martin, and W. Sun, A residual stiffness-based model for the fatigue damage of biological soft tissues. Journal of the Mechanics and Physics of Solids, 2020. 143: p. 104074.
38. Zhang, W. and M.S. Sacks, Modeling the response of exogenously crosslinked tissue to cyclic loading: The effects of permanent set. Journal of the mechanical behavior of biomedical materials, 2017. 75: p. 336-350.
39. Martin, C. and W. Sun, Modeling of long-term fatigue damage of soft tissue with stress softening and permanent set effects. Biomechanics and modeling in mechanobiology, 2013. 12: p. 645-655.
40. Henderson, B.S., K.F. Cudworth, E. Peña, and T.J. Lujan, Modeling fatigue failure in soft tissue using a visco-hyperelastic model with discontinuous damage. Journal of the Mechanical Behavior of Biomedical Materials, 2023: p. 105968.
41. Zitnay, J.L., G.S. Jung, A.H. Lin, Z. Qin, Y. Li, S.M. Yu, M.J. Buehler, and J.A. Weiss, Accumulation of collagen molecular unfolding is the mechanism of cyclic fatigue damage and failure in collagenous tissues. Science advances, 2020. 6(35): p. eaba2795.
42. Liu, D., S. Ma, H. Yuan, and B. Markert, Computational modelling of poro-visco-hyperelastic effects on time-dependent fatigue crack growth of hydrogels. International Journal of Plasticity, 2022. 155: p. 103307.
43. Rausch, M., W.D. Meador, J. Toaquiza-Tubon, O. Moreno-Flores, and A.B. Tepole, Biaxial mechanics of thermally denaturing skin-Part 2: Modeling. Acta biomaterialia, 2022. 140: p. 421-433.
44. Ateshian, G.A., Viscoelasticity using reactive constrained solid mixtures. Journal of biomechanics, 2015. 48(6): p. 941-947.
45. Zitnay, J.L., Y. Li, Z. Qin, B.H. San, B. Depalle, S.P. Reese, M.J. Buehler, S.M. Yu, and J.A. Weiss, Molecular level detection and localization of mechanical damage in collagen enabled by collagen hybridizing peptides. Nature communications, 2017. 8(1): p. 14913.
46. Dhume, R.Y., E.D. Shih, and V.H. Barocas, Multiscale model of fatigue of collagen gels. Biomechanics and modeling in mechanobiology, 2019. 18: p. 175-187.
47. Rodriguez, E.K., A. Hoger, and A.D. McCulloch, Stress-dependent finite growth in soft elastic tissues. Journal of biomechanics, 1994. 27(4): p. 455-467.
48. Han, T., T. Lee, J. Ledwon, E. Vaca, S. Turin, A. Kearney, A.K. Gosain, and A.B. Tepole, Bayesian calibration of a computational model of tissue expansion based on a porcine animal model. Acta Biomaterialia, 2022. 137: p. 136-146.
49. Blinkouskaya, Y. and J. Weickenmeier, Brain shape changes associated with cerebral atrophy in healthy aging and Alzheimer’s disease. Frontiers in Mechanical Engineering, 2021. 7: p. 705653.
50. Humphrey, J.D. and K. Rajagopal, A constrained mixture model for growth and remodeling of soft tissues. Mathematical models and methods in applied sciences, 2002. 12(03): p. 407-430.
51. Humphrey, J., Constrained mixture models of soft tissue growth and remodeling–twenty years after. Journal of Elasticity, 2021. 145(1): p. 49-75.
52. Zuo, D., S. Avril, H. Yang, S.J. Mousavi, K. Hackl, and Y. He, Three-dimensional numerical simulation of soft-tissue wound healing using constrained-mixture anisotropic hyperelasticity and gradient-enhanced damage mechanics. Journal of the Royal Society Interface, 2020. 17(162): p. 20190708.
53. He, Y., D. Zuo, K. Hackl, H. Yang, S.J. Mousavi, and S. Avril, Gradient-enhanced continuum models of healing in damaged soft tissues. Biomechanics and modeling in mechanobiology, 2019. 18: p. 1443-1460.
54. Zuo, D., Y. He, S. Avril, H. Yang, and K. Hackl, A thermodynamic framework for unified continuum models for the healing of damaged soft biological tissue. Journal of the Mechanics and Physics of Solids, 2022. 158: p. 104662.
55. Lee, T., M.A. Holland, J. Weickenmeier, A.K. Gosain, and A.B. Tepole, The geometry of incompatibility in growing soft tissues: Theory and numerical characterization. Journal of the Mechanics and Physics of Solids, 2021. 146: p. 104177.
56. Gacek, E., R.R. Mahutga, and V.H. Barocas, Hybrid discrete-continuum multiscale model of tissue growth and remodeling. Acta Biomaterialia, 2023. 163: p. 7-24.
57. Guo, Y., M.R. Mofrad, and A.B. Tepole, On modeling the multiscale mechanobiology of soft tissues: Challenges and progress. Biophysics Reviews, 2022. 3(3): p. 031303.
58. Sree, V.D. and A.B. Tepole, Computational systems mechanobiology of growth and remodeling: Integration of tissue mechanics and cell regulatory network dynamics. Current Opinion in Biomedical Engineering, 2020. 15: p. 75-80.
59. Estrada, A.C., K. Yoshida, J.J. Saucerman, and J.W. Holmes, A multiscale model of cardiac concentric hypertrophy incorporating both mechanical and hormonal drivers of growth. Biomechanics and modeling in mechanobiology, 2021. 20(1): p. 293-307.
60. Kitano, H., Systems biology: a brief overview. science, 2002. 295(5560): p. 1662-1664.
61. Chowkwale, M., M.L. Lindsey, and J.J. Saucerman, Intercellular model predicts mechanisms of inflammation‐fibrosis coupling after myocardial infarction. The Journal of Physiology, 2023. 601(13): p. 2635-2654.
62. Tan, P.M., K.S. Buchholz, J.H. Omens, A.D. McCulloch, and J.J. Saucerman, Predictive model identifies key network regulators of cardiomyocyte mechano-signaling. PLoS Computational Biology, 2017. 13(11): p. e1005854.
63. Irons, L., M. Latorre, and J.D. Humphrey, From transcript to tissue: multiscale modeling from cell signaling to matrix remodeling. Annals of biomedical engineering, 2021. 49(7): p. 1701-1715.
64. van Asten, J.G., M. Latorre, C. Karakaya, F.P. Baaijens, C.M. Sahlgren, T. Ristori, J.D. Humphrey, and S. Loerakker, A multiscale computational model of arterial growth and remodeling including Notch signaling. Biomechanics and Modeling in Mechanobiology, 2023: p. 1-20.
65. van Asten, J.G., T. Ristori, D.R. Nolan, C. Lally, F.P. Baaijens, C.M. Sahlgren, and S. Loerakker, Computational analysis of the role of mechanosensitive Notch signaling in arterial adaptation to hypertension. journal of the mechanical behavior of biomedical materials, 2022. 133: p. 105325.
66. Gierig, M., P. Wriggers, and M. Marino, Arterial tissues and their inflammatory response to collagen damage: A continuum in silico model coupling nonlinear mechanics, molecular pathways, and cell behavior. Computers in Biology and Medicine, 2023. 158: p. 106811.
67. Rahman, M.M., P.N. Watton, C.P. Neu, and D.M. Pierce, A chemo-mechano-biological modeling framework for cartilage evolving in health, disease, injury, and treatment. Computer methods and programs in biomedicine, 2023. 231: p. 107419.
68. Pensalfini, M. and A.B. Tepole, Mechano-biological and bio-mechanical pathways in cutaneous wound healing. bioRxiv, 2022.
69. Alon, U., Network motifs: theory and experimental approaches. Nature Reviews Genetics, 2007. 8(6): p. 450-461.
70. Milo, R., S. Shen-Orr, S. Itzkovitz, N. Kashtan, D. Chklovskii, and U. Alon, Network motifs: simple building blocks of complex networks. Science, 2002. 298(5594): p. 824-827.
71. Gilpin, W., Y. Huang, and D.B. Forger, Learning dynamics from large biological data sets: machine learning meets systems biology. Current Opinion in Systems Biology, 2020. 22: p. 1-7.
72. Liu, M., L. Liang, H. Dong, W. Sun, and R.L. Gleason, Constructing growth evolution laws of arteries via reinforcement learning. Journal of the Mechanics and Physics of Solids, 2022. 168: p. 105044.
73. Concha, F. and D.E. Hurtado, Upscaling the poroelastic behavior of the lung parenchyma: A finite-deformation micromechanical model. Journal of the Mechanics and Physics of Solids, 2020. 145: p. 104147.
74. Eskandari, M., T.M. Nordgren, and G.D. O’Connell, Mechanics of pulmonary airways: linking structure to function through constitutive modeling, biochemistry, and histology. Acta biomaterialia, 2019. 97: p. 513-523.
75. Roth, C.J., L. Yoshihara, and W.A. Wall, A simplified parametrised model for lung microstructures capable of mimicking realistic geometrical and mechanical properties. Computers in Biology and Medicine, 2017. 89: p. 104-114.
76. Comellas, E., S. Budday, J.-P. Pelteret, G.A. Holzapfel, and P. Steinmann, Modeling the porous and viscous responses of human brain tissue behavior. Computer Methods in Applied Mechanics and Engineering, 2020. 369: p. 113128.
77. Shi, L. and K. Myers, A finite porous-viscoelastic model capturing mechanical behavior of human cervix under multi-step spherical indentation. Journal of the Mechanical Behavior of Biomedical Materials, 2023. 143: p. 105875.
78. Conway, C.K., H.J. Qureshi, V.L. Morris, E.K. Danso, L. Desrosiers, L.R. Knoepp, C.J. Goergen, and K.S. Miller, Biaxial biomechanical properties of the nonpregnant murine cervix and uterus. Journal of biomechanics, 2019. 94: p. 39-48.
| Attachment | Size |
|---|---|
| fig_fatigue.png | 419.26 KB |
| fig_MS_mechanobiology.png | 231.38 KB |
| fig_ML.png | 658.85 KB |
| fig_springpot.png | 557.19 KB |
| fig_denature.png | 90.35 KB |
| fig_system_mech.png | 279.6 KB |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Beautifully done - Two topics missing (IMHO)
Beautifully done, my friend. I especially appreciate your focus on ML and "new" tissues. From an experimentalist's perspective, I'd like to add that constitutive models are only as good as the data with which - and procedures through which - they are informed. In my mind, areas that require future attention are: 1) better community buy-in to open data. Seeing folks make their well-curated and documented data openly available is still the exception. 2) experimental standards. No agreed-upon standards exist for how to test tissues and inform models. Together, these unresolved issues lead to a complete lack of reproducibility, and if we are serious about building reliable and predictive constitutive models, both need to be addressed (in addition to the issues you have been raising). I look forward to doing that with you :).
In reply to Beautifully done - Two topics missing (IMHO) by manuel.rausch@…
yes, next journal club we can do testing standards?
Great points, both very true.
1) How can we increase this buy-in? I think computational models are more widely shared through Github, but it is true that experimental data sharing is not always as widely available. Maybe we should start a page with links to multiple datasets? Is there something like that already?
2) This definitely sounds like a topic for a journal club. Quick thought, even while we converge to a standard, we should be able to relate data across different studies of the same tissue if we know boundary conditions. Not ideal, but at least we can address some of the issues with 'direct comparison' when such direct comparison is not possible. How much of the lack of reproducibility is due to actually getting data that is not compatible between tests, and how much of it is 'compatible data' but only once you can account for boundary/loading conditions? And if it is in principle possible to relate data from different test modes, is the issue not just that the protocol is not standardized but that it is not reported in necessary detail? If it is not possible to relate data across studies even accounting for boundary conditions, then what is the issue? Tissue processing? Personally, the thought of trying to piece together data across many different studies with different protocols by using the same constitutive model but trying to fill in the different boundary conditions (with uncertainties) sounds very interesting. Ultimately, though, agree what we need is a standard!