
Mechanics of living mammalian cytoplasm
1.Overview
The cytoplasm of living mammalian cells is a crowded, yet dynamic environment(1). It provides the key physical environment to the cellular factory and all intracellular physiological processes from transcription, translation, to protein binding and folding. Therefore, understanding the fundamental physical nature of the cytoplasm is critical to understanding the basic physiology of cells. Moreover, there are continuous intracellular movements that are vital for cell function, such as transport of vesicles and other organelles. While biological motors and other enzymatic processes provide key driving forces for these activities, the mechanical behavior of the cytoplasm are crucial for determining the mechanical resistance that cellular compartments experience. Both the active driving force and appropriate mechanical environment are critical for shaping the living cellular machinery. However, while the force that molecular motors generate both individually and collectively has been extensively studied(2, 3), the mechanical behavior of the cytoplasmic environment remains largely explored.
Indeed, it has become increasingly recognized that the mechanical properties of cells have direct impact on biological functions of cells and tissues(4-13); form example, the stiffness of cells and their surrounding matrix influence stem cell fate, cancer metastasis, apoptosis, cell signaling, etc. Nevertheless, most of previous studies characterizing cell mechanics probed cells from exterior, using techniques such as atomic force microscopy, magnetic tweezers, or optical stretcher(14-20); thus these measurements were rather mainly determined by the mechanics of the stiff actin-rich cell cortex than the much-weaker and softer cytoplasm(21). Moreover, previous mechanical characterization of cells and the cytoplasm were mostly tested within small deformation regime(3-5, 22-24), while cells often undergo extremely large deformation or experience repeated cycles of loading and unloading during numerous physiological processes, such as cell migration, tumor cell extravasation, and intracellular transport; thus the nonlinear mechanical behavior of living mammalian cytoplasm at large deformation remains largely unexplored.
My group has been using optical tweezers to perform active microrheology measurement(3, 21, 23, 24) to directly characterize mechanical behaviors of living mammalian cytoplasm. Other techniques such as magnetic tweezers are also useful in studying cytoplasmic mechanics. Here I would like to discuss a few topics that may provide a comprehensive picture of the cytoplasm of living mammalian cells, from a mechanics and materials standpoint. I will continue modify and expand interactively.
2.1 Cytoplasmic mechanics at the linear regime.
The cytoplasm has been commonly revealed as a viscoelastic material, whose property does not depend on probe size(25). Recently, it has also been suggested that the cytoplasm of living cells may behave like a poroelastic gel, whose property instead strongly depends on the length scale of the probe(19). However, in contrast to these elasticity-based models, there were also studies claiming that the cytoplasm of living mammalian cells behaves like a viscous fluid, rather than an elastic solid, and the viscosity of the cytoplasm is of several hundred times greater than that of water(26). These understandings are remarkably distinct. Therefore, a systematic characterization of the cytoplasmic mechanical properties, even in the small deformation regime, across a wide strain rate and size range would help us understand where the different interpretations might come from. Such characterization will provide important insights into understanding the physical environment and numerous key dynamic processes inside living cells.
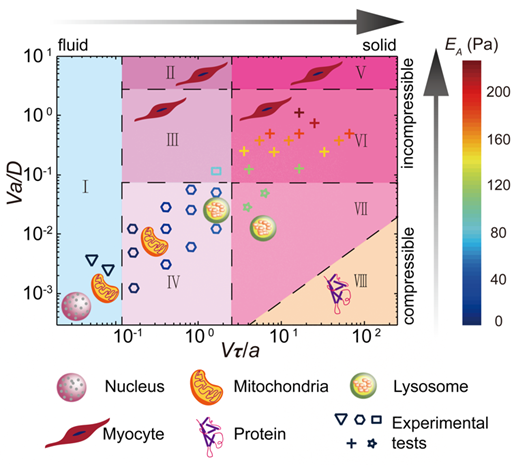
In a recent study(24), we use optical tweezers to drag a plastic bead in the cytoplasm of a living mammalian cell, and directly measure the force and displacement relationship, which reflects the mechanical behavior of the cytoplasm. With considering both viscoelasticity and poroelasticity, we identify two independent dimensional parameters in the experiments: V/a and Va, where V and a represent the speed and diameter of the probe bead. Using these two control parameters, and through a combination of experimental measurement and scaling analysis, we reveal different origins of cytoplasmic resistance, ranging from viscous, viscoelastic, poroelastic, to pure elastic. Finally, we classify different cytoplasmic mechanical behaviors in an overall state diagram, which illustrates different origins of mechanical resistance of a variety of physiological processes in cells with different characteristic size and speed(24).
Effects of cytoskeletal components. The following up question is to investigate the contribution of major cytoplasmic components to the mechanical behaviors of the cytoplasm. To do so, chemicals and genetic tools can be used to disrupt or knock out specific cytoskeletal components; moreover, cellular water amount can be also controlled through application of external osmotic pressure. For example, cytochalasin D and nocodazole can be used to depolymerize F-actin and microtubules (MTs). Cytoplasmic intermediate filament (IF), such as vimentin IF, can be genetically knocked out while maintaining the same amount of actin and microtubules(27). Our previous results have shown that vimentin IFs indeed greatly impact mechanics of the cytoplasm(21). It is critical to characterize the impact of major cytoplasmic components, such as F-actin, MTs, vimentin IFs, water content, and motor activity, to the cytoplasmic mechanical behaviors. This will also help us understand the role of each cytoskeletal component in determining the cytoplasmic viscoelasticity, poroelasticity, and viscosity.
2.2 Nonlinear mechanical behavior of mammalian cytoplasm at large deformation.
Cells often experience large deformation during the development, migration, long distance active transport and other important life processes(1). At large deformation, biomaterials often exhibit nonlinear behavior, whose modulus changes as a function of strain(28). This behavior has been well studied for reconstituted biopolymer networks in vitro(29, 30). However, most studies on living cells rather focus on probing their property at small deformations, measuring the stiffness of the cell, which corresponds to the linear elastic modulus(3, 14, 21, 31-33). Yet, the nonlinear mechanical behavior of the cytoplasm at large deformation remains largely unknown; moreover, to the extreme case, the yield strength inside living cells has never been measured. A direct characterization of the yielding behavior of the cytoplasm is of particular importance, as it provides crucial insights into the cell’s ability to maintain mechanical integrity under complex physiological conditions, and will also provide key evidence for the glassy material model recently proposed to describe the cytoplasm(34). In this Aim, we plan to directly measure the nonlinear mechanical behavior of living mammalian cytoplasm. We will further obtain the yield strength/strain and post-yielding behavior of the cytoplasm. Furthermore, we will measure the contribution of each major cellular component to the nonlinear mechanical behaviors of the cytoplasm and key mechanical parameters. These would enable a quantitative understanding of cell properties and behaviors in the nonlinear regimes.
Yield strength/strain and post-yielding behavior of cytoplasm. The mesh size of typical cytoskeletal network in mammalian cells is about 50 nm(35), however most endogenous organelles or vesicles are much larger than it. Thus, to enable long distance transport from one side of a cell to the other side, the active force applied onto the cargo or organelle must be larger than the mechanical yield strength of the cytoplasm. Because of the complex microstructure of the cytoplasm, yield strength/strain and post-yielding behavior are expected to have pronounced size and rate-dependence.
Effects of cytoskeletal components. It is entirely unclear how each of the cytoskeletal components determines the nonlinear mechanical behaviors of the cytoplasm at large deformations. In particular, vimentin IFs have been shown to play an important role in many key physiological processes such as epithelial to mesenchymal transition (EMT) and wound healing. During these processes, the expression of vimentin IFs is strongly associated with the ability of cells to deform and migrate(27, 36, 37). Thus, the role of IFs in the nonlinear mechanics of the cytoplasm may be more important.
2.3 Mechanical behavior of mammalian cytoplasm with cyclic loading-unloading.
During numerous physiological processes, such as cell migration and division, tumor invasion, wound healing, cells do not just simply deform moderately or to a large extent, they also undergo these deformations repeatedly as part of their “daily life”. This highlights the dynamic nature of the cell cytoplasm as a living material. Characterizing plasticity, shape recovery, damage and self-healing capability of the cytoplasm under repeated loading is critical for understanding the fundamental cell physiology, yet remains largely unexplored. These mechanical behaviors are crucial for the applications of most engineering materials, however, have never been probed in the cytoplasm, therefore limit our understanding of the mechanical environment inside living cells. Characterization of these complex mechanical parameters of the cytoplasm under cyclic loading will also provide key insights into understanding how cells utilize different cellular components to assemble living machinery and will motivate the designs of engineering living systems.
2.4 plasticity
2.5 self-healing properties
Figure 1. Mechanical state diagram of the cytoplasm. Symbols label our preliminarily tested states. I: viscous fluid; II: incompressible viscoelasticity; III: poro-viscoelasticity; IV: compressible viscoelasticity; V: incompressible elasticity; VI: unrelaxed poroelasticity; VII: compressible elasticity; VIII: continuous mechanics not applicable.
Figure 2 Mechanical state diagram of the cytoplasm. Symbols label our preliminarily tested states. I: viscous fluid; II: incompressible viscoelasticity; III: poro-viscoelasticity; IV: compressible viscoelasticity; V: incompressible elasticity; VI: unrelaxed poroelasticity; VII: compressible elasticity; VIII: continuous mechanics not applicable.
Bibliography
1.B. Alberts, Molecular biology of the cell. (2015).
2.A. G. Hendricks, E. L. F. Holzbaur, Y. E. Goldman, Force measurements on cargoes in living cells reveal collective dynamics of microtubule motors. Proceedings of the National Academy of Sciences 109, 18447-18452 (2012).
3.M. Guo et al., Probing the Stochastic, Motor-Driven Properties of the Cytoplasm Using Force Spectrum Microscopy. Cell 158, 822-832 (2014).
4.P. A. Janmey, C. A. McCulloch, Cell Mechanics: Integrating Cell Responses to Mechanical Stimuli. Annual Review of Biomedical Engineering 9, 1-34 (2007).
5.D. A. Fletcher, R. D. Mullins, Cell mechanics and the cytoskeleton. Nature 463, 485-492 (2010).
6.D. E. Discher, P. Janmey, Y. L. Wang, Tissue cells feel and respond to the stiffness of their substrate. Science 310, 1139-1143 (2005).
7.A. J. Engler, S. Sen, H. L. Sweeney, D. E. Discher, Matrix elasticity directs stem cell lineage specification. Cell 126, 677-689 (2006).
8.O. Chaudhuri et al., Extracellular matrix stiffness and composition jointly regulate the induction of malignant phenotypes in mammary epithelium. Nat Mater 13, 970-978 (2014).
9.N. Huebsch et al., Harnessing traction-mediated manipulation of the cell/matrix interface to control stem-cell fate. Nature Materials 9, 518-526 (2010).
10.S. Huang, D. E. Ingber, Cell tension, matrix mechanics, and cancer development. Cancer Cell 8, 175-176 (2005).
11.M. J. Paszek et al., Tensional homeostasis and the malignant phenotype. Cancer Cell 8, 241-254 (2005).
12.J. Fu et al., Mechanical regulation of cell function with geometrically modulated elastomeric substrates. Nature Methods 7, 733-U795 (2010).
13.R. J. Pelham, Y. L. Wang, Cell locomotion and focal adhesions are regulated by substrate flexibility. Proc. Natl. Acad. Sci. U. S. A. 94, 13661-13665 (1997).
14.S. Y. Tee, J. P. Fu, C. S. Chen, P. A. Janmey, Cell Shape and Substrate Rigidity Both Regulate Cell Stiffness. Biophysical Journal 100, L25-L27 (2011).
15.J. Guck et al., The optical stretcher: a novel laser tool to micromanipulate cells. Biophysical Journal 81, 767-784 (2001).
16.J. M. Maloney et al., Mesenchymal Stem Cell Mechanics from the Attached to the Suspended State. Biophysical Journal 99, 2479-2487 (2010).
17.J. Solon, I. Levental, K. Sengupta, P. C. Georges, P. A. Janmey, Fibroblast Adaptation and Stiffness Matching to Soft Elastic Substrates. Biophysical Journal 93, 4453-4461 (2007).
18.B. Fabry et al., Scaling the microrheology of living cells. Phys Rev Lett 87, 148102 (2001).
19.E. Moeendarbary et al., The cytoplasm of living cells behaves as a poroelastic material. Nat Mater 12, 253-261 (2013).
20.Hadi T. Nia, L. Han, Y. Li, C. Ortiz, A. Grodzinsky, Poroelasticity of Cartilage at the Nanoscale. Biophysical Journal 101, 2304-2313 (2011).
21.M. Guo et al., The Role of Vimentin Intermediate Filaments in Cortical and Cytoplasmic Mechanics. Biophysical Journal 105, 1562-1568 (2013).
22.G. Bao, S. Suresh, Cell and molecular mechanics of biological materials. Nature materials 2, 715 (2003).
23.S. K. Gupta, M. Guo, Equilibrium and out-of-equilibrium mechanics of living mammalian cytoplasm. Journal of the Mechanics and Physics of Solids, (2017).
24.J. Hu et al., Size- and speed-dependent mechanical behavior in living mammalian cytoplasm. Proceedings of the National Academy of Sciences, (2017).
25.P. Kollmannsberger, B. Fabry, Linear and Nonlinear Rheology of Living Cells. Annual Review of Materials Research 41, 75-97 (2011).
26.J. F. Berret, Local viscoelasticity of living cells measured by rotational magnetic spectroscopy. Nature communications. 2016 (10.1038/ncomms10134).
27.M. G. Mendez, S.-I. Kojima, R. D. Goldman, Vimentin induces changes in cell shape, motility, and adhesion during the epithelial to mesenchymal transition. The FASEB Journal 24, 1838-1851 (2010).
28.C. Storm, J. J. Pastore, F. C. MacKintosh, T. C. Lubensky, P. A. Janmey, Nonlinear elasticity in biological gels. Nature 435, 191-194 (2005).
29.M. L. Gardel et al., Prestressed F-actin networks cross-linked by hinged filamins replicate mechanical properties of cells. Proc. Natl. Acad. Sci. U. S. A. 103, 1762-1767 (2006).
30.Y.-C. Lin et al., Origins of Elasticity in Intermediate Filament Networks. Physical Review Letters 104, 058101 (2010).
31.J. C. del Álamo, G. N. Norwich, Y.-s. J. Li, J. C. Lasheras, S. Chien, Anisotropic rheology and directional mechanotransduction in vascular endothelial cells. Proceedings of the National Academy of Sciences 105, 15411-15416 (2008).
32.Y. Tseng, T. P. Kole, D. Wirtz, Micromechanical mapping of live cells by multiple-particle-tracking microrheology. Biophysical Journal 83, 3162-3176 (2002).
33.A. J. Engler, S. Sen, H. L. Sweeney, D. E. Discher, Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 126, 677-689 (2006).
34.Bradley R. Parry et al., The Bacterial Cytoplasm Has Glass-like Properties and Is Fluidized by Metabolic Activity. Cell 156, 183-194 (2014).
35.K. Luby-Phelps, Cytoarchitecture and physical properties of cytoplasm: Volume, viscosity. diffusion, intracellular surface area. International Review of Cytology - a Survey of Cell Biology, Vol 192 192, 189-221 (2000).
36.I. Y. Wong et al., Collective and individual migration following the epithelial–mesenchymal transition. Nat Mater 13, 1063-1071 (2014).
37.M. G. Mendez, D. Restle, P. A. Janmey, Vimentin Enhances Cell Elastic Behavior and Protects against Compressive Stress. Biophysical Journal 107, 314-323 (2014).
| Attachment | Size |
|---|---|
| cytoskeletal structures.png | 850.13 KB |
| Picture1.png | 139.35 KB |
{kind=link}
{kind=link}
Modelling combinations of mechanics and biological function
<p>Dear Guoming,</p>
<p>This is a very interesting topic. Are you aware of any studies that looks at multiscale computational modeling of changes in the cellular function (protein expression, diffusion, ion gating, etc) given changes in the cells' mechanical environment?</p>