Mechanobiology and Mechanomedicine: tuning the tension in the life
Baohua Ji (bhji [at] zju.edu.cn)
Department of Engineering Mechanics, Zhejiang University, Hangzhou 310027
Abstract
Mechanobiology is now a widely accepted field of science at the interface of biology, medicine, engineering, and physics. It focuses on how the physical forces and changes in the mechanical properties of cells and tissues contribute to development, cell differentiation, physiology, and disease. Mechanomedicine, however, is an emerging field of diagnosing and treating diseases based on the knowledge obtained from mechanobiological studies. It proposes a new concept of diagnosis and treatment of diseases using the mechanical concept, theories, and approaches. But the question how we employ the mechanical factors to diagnose and treat the diseases is far from being addressed. Here, based on our recent studies on tensional homeostasis, we try to give some clues for the endeavor of answering these questions. Diseases are thought as the results of significant deviation of function/structure of life from the corresponding homeostasis at different length scales, including the tensional homeostasis. If we can properly tune the value of tension in the living organisms, then it creates a new way of treating the diseases.
1. Introduction
The term ‘mechanobiology’ has now been widely accepted. In this prosperous field, broad research approaches, ranging from basic biology to medical research, have been conducted from the perspective of mechanobiology. Compared to mechanobiology, ‘mechanomedicine’ is relatively new. The term of ‘mechanomedicine’ first appeared in the paper by Dr. Bloomfield in 1955 [1]. But the concept and connotation of mechanomedicine is firstly summarized by Dr. Ning Wang in 2009 in iMechanica Journal club (https://imechanica.org/node/5102). In a recent personal communication, he further enriched the concept of the mechanomedicine as the mechanobiology- and biomechanics- based medicine, which is a contrast to the chemical-based medicine and may offer a new way of diagnosing and treating the diseases. In fact, the application of biomechanics/mechanobiology to medicine and health has actually existed for a very long time.
But previous applications of biomechanics/mechanobiology in the treatment/diagnosis of diseases are limited in some specific areas, and had not been guided in a quantitative way because the underlying mechanisms have not been understood. For instance, why can we tune the physiology and pathology of life through the mechanical approaches? And how should we tune it? These questions have not been addressed.
According to the definition in the textbook of pathology, a disease means a condition marked by pronounced deviation from the normal healthy state [2]. It causes distinct changes in tissues and cells in an individual, named as lesion. The mission of medicine is to repair/tune back these injuries to the normal states. These normal states are called homeostatic states. The organisms have the ability to keep the homeostatic states by which they keep internal conditions near constant to function efficiently. However, the homeostasis usually indicates the chemical and thermal factors, such as the temperature, blood PH, glucose concentration, etc. Recent studies suggested that deviation from mechanical homeostasis is surely associated with diseases, including aberrant wound repair, developmental abnormalities and cancer [3]. But we know little about the mechanical aspects of the homeostasis.
2. The tensional homeostasis
2.1 The tension at different scales
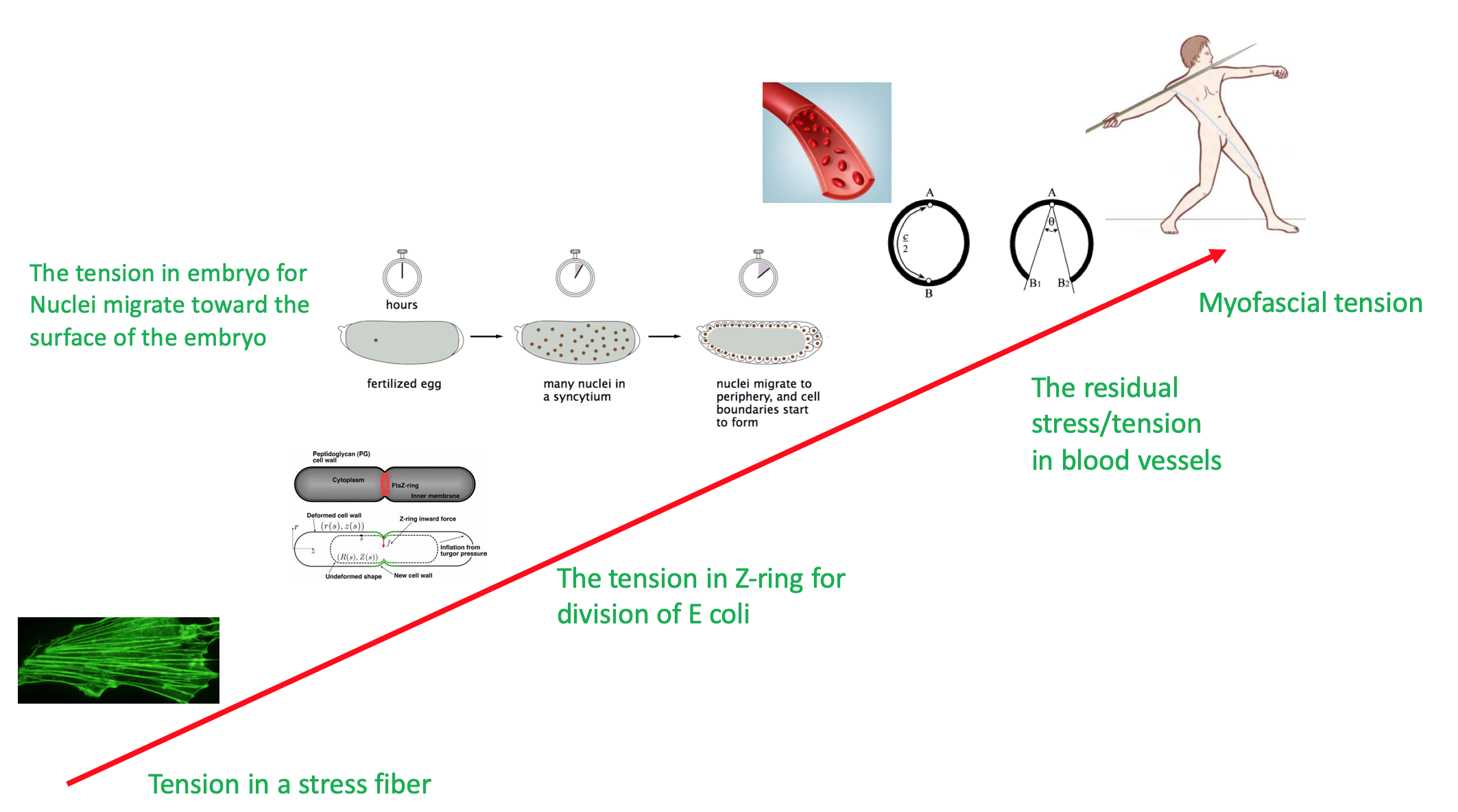
The tension universally presents in the living organisms of different scales. At the cellular and sub-cellular scales, a typical example is the tension force in the actin cytoskeleton that is produced by acto-myosin-II, which plays crucial roles in cell mechanosensing behaviors [4]. A similar structure is found in the Z-ring [5] that forms during the division of E coli that produces a large tension force dividing the cell into two separate parts (Figure 1). The intra-cellular tension force can also be produced by the microtubules for cell mitosis [6].
Figure 1. Typical tension forces present at different scales in the living organisms.
At the multi-cellular level, for instance, in the early Drosophila embryogenesis, there is tension in the embryo driving thousands of nuclei migrating toward the surface of the embryo, by which a layer of follicle cells forms at the surface of the embryo [7]. During the process, actomyosin contractility induces contractions of the cortex which generates a large-scale flow of the cytoplasm, which churns the cytoplasm and moves the nuclei along the anteroposterior axis, bringing them closer to the surface. In addition, during dorsal closure of the embryogenesis of Drosophila, the amnioserosa monolayer contracts actively with the assistance of a supracellular actin cable along the monolayer boundary, to pull the two lateral epidermal sheets move dorsally to cover the amnioserosa and ultimately fuse at the dorsal midline [8].
At the macroscopic level, there exhibits notable tension in the tissue as well. A well-known example is the residual stress/tension in blood vessels, which has been systematically studied by Y-C Fung [9]. His studies showed that the residual strain and stress in the blood vessels could produce large opening angles when the blood vessels are cut along the longitudinal axis of the vessel. This residual stress is crucial for distributing the stress uniformly in the tissue and remodeling the blood vessel.
At even larger scale than tissues, the anatomy shows that the muscles and their accompanying fascia form body size scale soft skeleton structures along the body, called myofascial meridians. The tension of these elastic myofascial chains will help to carry out any kind of functional movement of the whole body.
2.2 The crucial roles of tension in organisms
The tension takes crucial roles in the biological functions of living organisms at different scales. For instance, Wang and Suo [10] had shown that the pretension in the cytoskeleton is crucial for long distance force propagation. The characteristic length of the force transfer is proportional to the square root of the prestress in the cytoskeleton. Recent studies showed that tuning the tension in the cytoskeleton with mechanical and chemical approaches can regulate the deformation of the nucleus and thus the degree of gene expression [11].
The tension force is also important for neuronal signal transmission. Siechen et al [12] showed that when the tension in the axon was relaxed by cutting the axon with laser, the accumulation of vesicles at the pre-synaptic membrane was retarded. But once the tension force on the axon was recovered by using a MEMS loading device, the accumulation of the vesicle got back. In addition, the fusion of the vesicles with the pre-synaptic membrane depends on the tension force applied by the fusion proteins [13].
In the alveolus of our lung, the tension in the alveolus membrane is for producing the pressure for balancing the air pressure in the alveolus. When the pneumocytes was attacked, e.g., by the COVID-19 virus, the pneumocytes will not be able to secrete surfactant, which will cause a dramatic increase of the membrane tension in the alveolus, which then consequently induce the collapse of the alveolus, and thus cause the patients being short of breath.
Recent studies showed that the tension force also takes crucial roles in the regulation of the glycolysis of cells [14]. For instance, it is found that higher rates of glycolysis are related to the presence of thicker F-actin structures, i.e., the glycolysis depends on the magnitude of the tension force in the cytoskeleton which modulates the activity of the mitochondrion.
2.3 Typical tensional homeostatic values in living organisms
Experimental observations suggested the magnitude of the tension at different scales may have homeostatic values. For instance, the pre-strain in stress fibers is 0.1–0.2 [4,15]. Correspondingly, the tension force applied on focal adhesion by the stress fiber produces a homeostatic value of shear stress around 5kPa [16,17]. At the cellular level, the pre-strain is approximately 0.1 under physiological conditions [15,18]. At the larger scales, there exist homeostatic pre-strains in tissues as well, such as bone, tendon, or blood vessel [19-21]. However, the underlying mechanisms that determine the value of the homeostatic pre-strain in cells and tissues remain to be clarified.
Bias from the above tensional homeostatic values may induce injuries of cells, tissue and organ. For instance, the bias of the shear stress on focal adhesion from the homeostatic value will cause the loss of the stability of the focal adhesion, which then impairs the viability of cells. Correspondingly, when the mechanical stress felt by bone cells is biased from its homeostatic value, abnormal bone remodeling will be activated and leads to bone diseases, such as the bone loss of astronauts under microgravity conditions in outer space.
What is the mechanical function of the tensional homeostasis? How does it enable cells to be mechanosensitive and how does this behavior propagates to larger scales? In the following, we will discuss these problems through a few examples.
3. From the tension homeostasis to the ‘structural’ homeostasis
3.1 Traction-distance law
For cell adhering to the substrate, interaction force between cell and matrix, called traction force, will be generated at the cell-matrix interface because of the contractility of cytoskeleton. That is, the tension force produced by the contractility of cytoskeleton is balanced by the traction force from the substrate in the cell-matrix system. The traction force will produce deformation on the substrate underneath the cells [22]. Therefore, one can calculate the traction force by measuring the deformation of the substrate.
Some typical observations on the distribution of the traction force are eminently attractive [23]: The traction force is larger at the edge of the cell than that at the center of the cell; The traction force is higher for the large spreading area than for the small one; The distribution of the traction force is closely correlated to the distribution of the size of focal adhesion: the focal adhesion near the edge is larger than that in the inner region of the cell, and correspondingly it is large for cell with the large spreading area than for cell with the small spreading area. However, the underlying mechanisms of the distribution of traction force and size of focal adhesion at the cell-matrix interface are not known before.
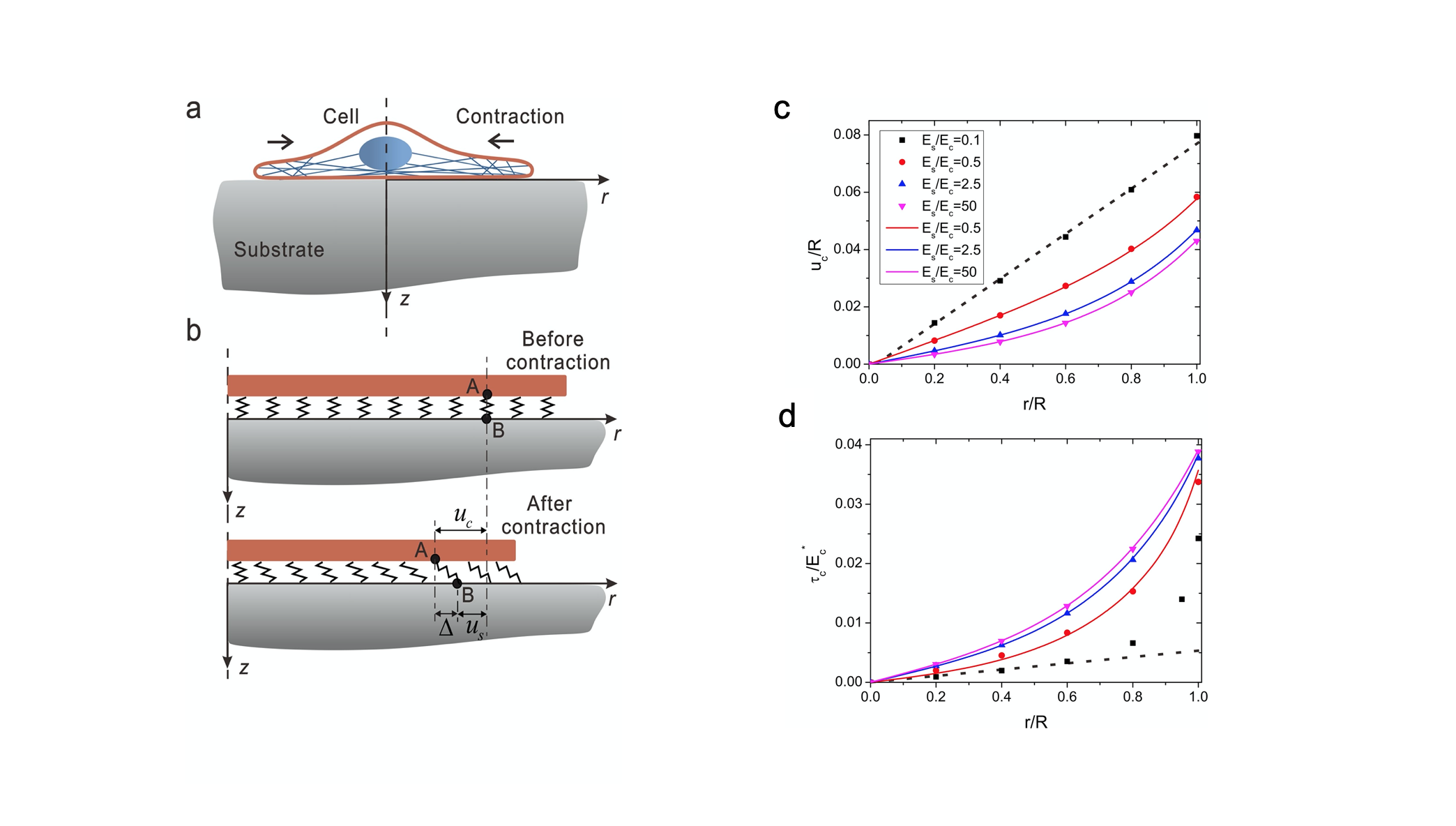
A theoretical model was developed for analyzing the distribution of the traction force [24]. For simplicity, they model the cell by a self-contract elastic disc, to consider the pre-strain in the cell due to the contractility of cytoskeleton. The substrate is modeled as a semi-infinite elastic body. And between the cell and matrix, there is a layer of continuously distributed elastic spring for considering the adhesion of cell to the substrate (Figure 2). Based on this model, a perturbation solution of the traction force was obtained, showing that the traction force is increasing with the distance from the cell center, which is named as ‘Traction-distance law’. This law is primary for understanding the mechanosensing behaviors of cells, which will be discussed as follows.
Figure 2. The contracting-disk model of cell–matrix interaction. (a) Schematic of a cell adhering to and pulling on an elastic substrate, owing to the intrinsic contractility of the cell. (b) The cell is modeled as an elastic contracting disk that is anchored to the substrate via molecular bonds. (c) The cell displacement increases with distance from the cell center. (d) The traction increases with distance from the cell center [24].
3.2 Revisit of the cell polarization
The concept of cell polarization should be revisited based on the finding of traction-distance law. According to Wikipedia, ‘cell polarity’ refers to the intrinsic asymmetry observed in cells, either in their shape, structure, or organization of cellular components. As we can see, this definition is made only from the point of view of chemistry and structure. In fact, besides the structural change, the mechanical aspects of change in cell polarization should be included.
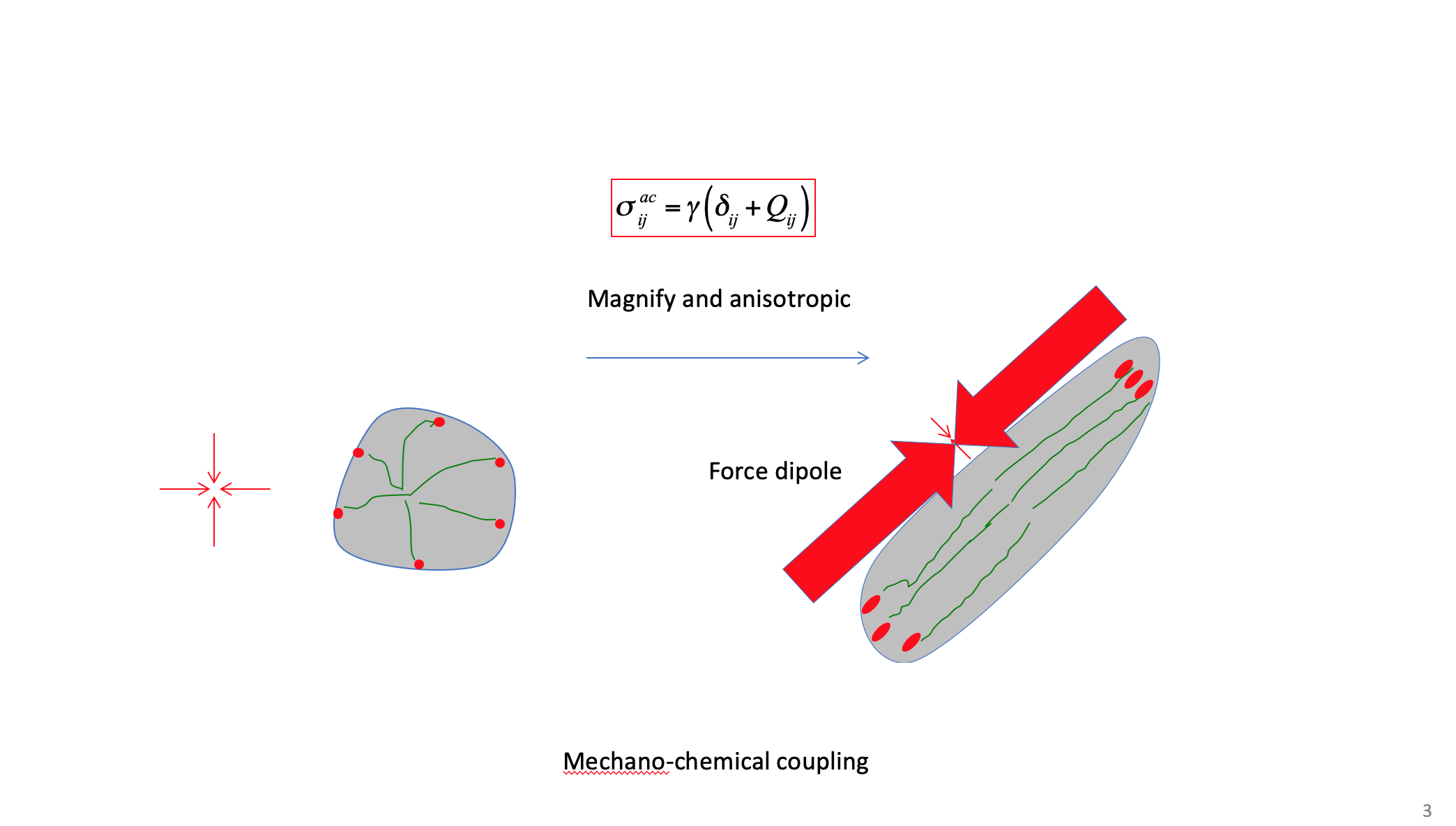
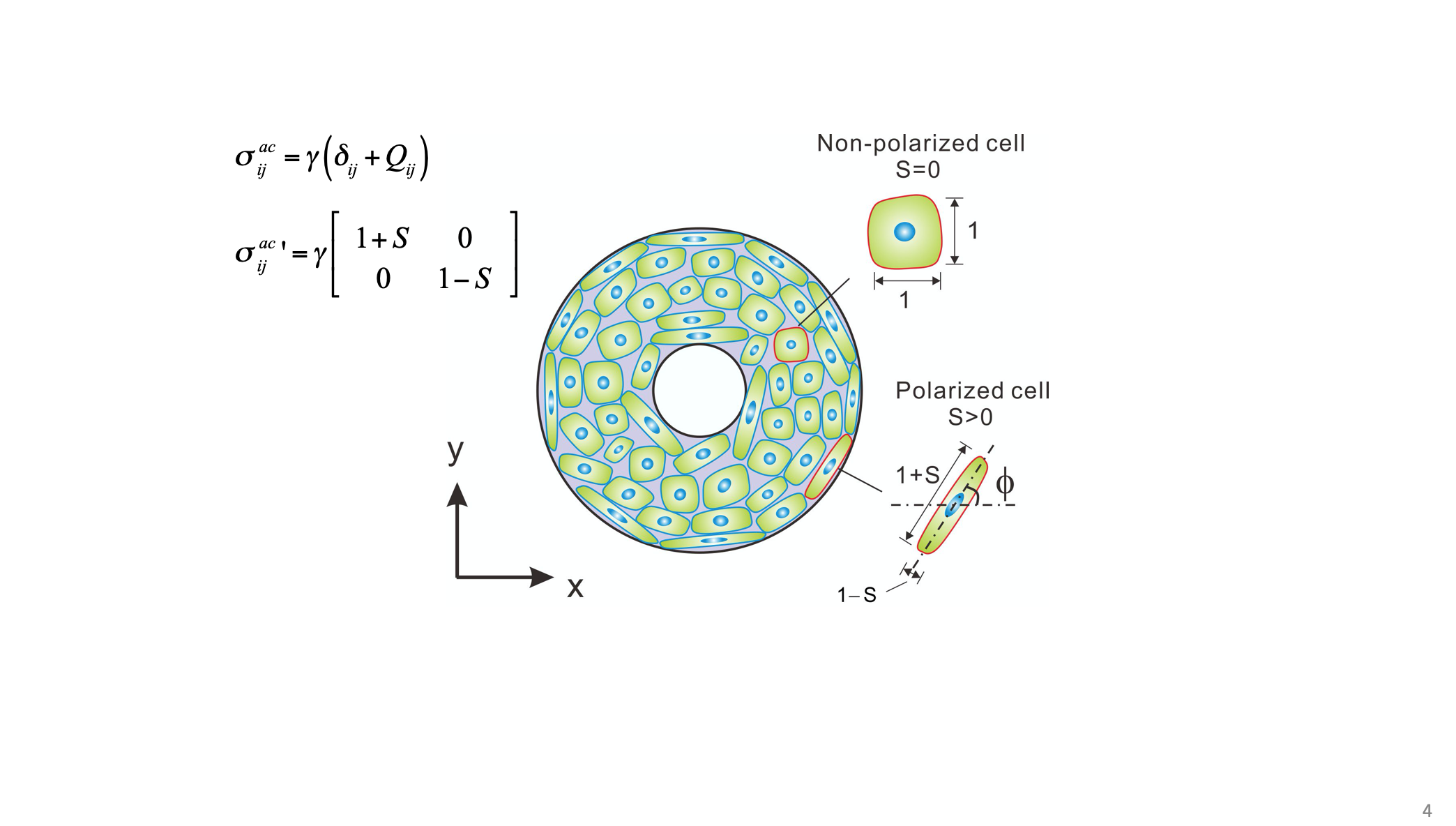
According to Figure 3, the typical structural features of cell polarity are areal and geometric change of cell (an increase of area and aspect ratio), and the associated polymerization and re-arrangement of cytoskeleton, as well as the areal and geometric change of focal adhesion (an increase of area and aspect ratio). According to the traction-distance law, the tension force by cytoskeleton along the long axis of the cell should be larger than that along the short axis of the cell. That is, not only the structure, but also the tension force and its distribution are changed. This is a new understanding of cell polarization from the point of view of physics/ mechanics. The mechanical character of cell polarization can be expressed by a tensor, called cell active stress, σ_ij^ac=γ(δ_ij+Q_ij), where Q_ij is the nematic order parameter, which describes the anisotropy of the active stress due to the polarity of cell [25].
"Q"=(■(p&q@q&-p))
When Q_ij is equal to zero, the active stress is isotropic (corresponding to un-polarized cell), otherwise, it is anisotropic (corresponding to the polarized cell). The principal active stress, i.e., the stress along the axes of the cell, can be expressed as
σ_ij^(ac ')=γ(■(1+S&0@0&1-S))
According to the traction-distance law, the aspect ratio of the polarized cell is then estimated by α=(1+S)/(1-S). In this context, cell polarization shows a typical coupling between the chemistry/structure and the mechanics at the cellular level in organisms.
Figure 3. The mechanical view of cell polarization and revisit of the ‘old’ definition of cell polarization which only considers the structural change in the polarization.
3.3 Production of the driving force for cell migration
It is found that there is a biphasic dependence of the stability of focal adhesion on the traction force: when the force is smaller than a critical value, the focal adhesion nucleates and grows; when the force is larger than the critical value, it disassembles [17]. This biphasic response of focal adhesion is smartly used by the cell for regulating the formation of focal adhesion at the front of the cell, and the disassembly of focal adhesion at the rear during cell migration [23,26].
We find that the regulation relies on a genius combination of the biphasic force dependence of focal adhesion and the distribution of traction at the cell-matrix interface controlled by the traction-distance law. Because of the polarized shape, the traction is small at the cell front, then the traction promotes the nucleation and growth of focal adhesion, which enables the cell body to move forwards with the help of the firm adhesion at the cell front; while at the cell rear, the traction is so large that the traction disassembles the focal adhesion for detaching of the cell rear. That is, the cell can regulate the stability of focal adhesion at any given location by modulating its spreading shape. In this way, the cell is able to control the speed and direction of its migration by its polarized shape.
3.4 Stress-driven cell polarization and alignment
For multiple-cell system, such as the 2D cell layer, we developed the continuum model for studying the collective behaviors of cells [25]. According to the chemical-mechanical coupling of cell polarization, the active stress of the cell should be considered in the force balance of the system, correspondingly the energy of cell polarization and alignment should be included in the free energy of the system. These considerations built a new cell model, named as nematic cell model for analyzing the multiple cell behaviors in a 2D cell layer (Figure 4). The force balance equation is given by
h_c (σ_(ij,j)^el+σ_(ij,j)^ac )-ρk_eff u_i=0
Since σ_(ij,j)^ac as the function of Q_ij was included in the equation, more relations should be introduced for solving the problem.
For this purpose, the energy of cell polarization and alignment were considered in the free energy of the system as
F=h_c ∬_Ω▒[α/2 S^2+k/2 Q_(ij,j)^2-ηQ_ij ε_ij+f_el (ε_ij)]dA
Minimizing the free energy results in the additional equations for Q_ij, which enable us to solve the problem and predict the polarization and orientation of cells.
Figure 4. Schematic illustration of the nematic cell model considering the cell polarization and orientation [25].
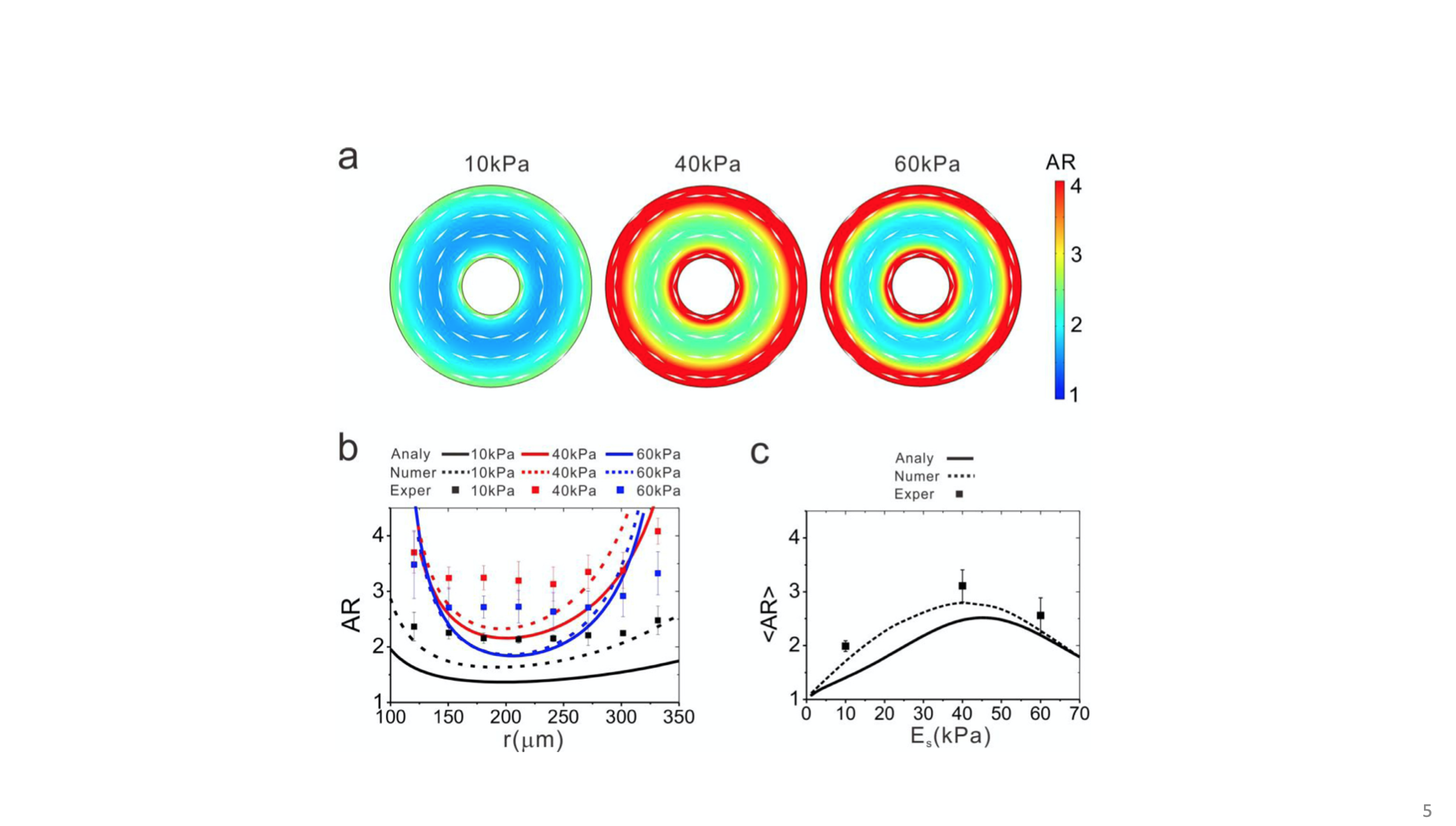
For the ring-like pattern, the analytical solution can be obtained of which the predictions of the orientation and polarization of cells agree well with the experiments. The results suggested that the cell polarization prefers to be aligning with the direction of the maximum principal stress (by this the cells consume the minimum energy to develop necessary homeostatic tension in the cell). For instance, with the traction free boundary condition in the ring-like cell layer, the maximum principal stress is predicted to be along the circular direction, consistent with the experimental results that the cells align along the circular direction. Imagining that if the cell is not aligning with the principal stress, then the cytoskeleton will bear the shear stress which shifts them toward the direction of the principal stresses that with zero shear stress.
Figure 5. Cell polarization and alignment on a ring pattern of different substrate stiffness. (a) The model prediction. The color map illustrates the cell aspect ratio (AR) and the white spindles indicate the cell orientation. (b) Cell AR as function of the radial coordinate for different substrate stiffness. (c) The mean value of cell aspect ratio biphasically depends on the substrate stiffness [25].
Secondly, the theoretical predictions show that the aspect ratio of cells at the edge of the ring is larger than that in the inner region of the ring. Correspondingly, at the edge of the ring, there is larger in-plane maximum shear stress than that in the inner region. These results seem to suggest that the polarization of cells is driven by the in-plane maximum shear stress, as the cytoskeleton barely sustains shear stress [27]. In addition, the maximum shear stress is an indicator of the anisotropy of the in-plane stresses. And the magnitude of the maximum shear stress represents the degree of the anisotropy of the in-plane stresses and thus of the cell polarity.
If there is traction at the boundary, the orientation of cells will be different. In the wound healing process, for example, there is formation of actin ring at the edge of the wound. Due to the contraction of the actin ring, there develops tension force at the wound edge, which equivalently produces pressure around the edge. By considering the effect of the actin ring in the boundary condition, the model predicted that the cell orientation changed from being along the circular direction to being along the radial direction because the maximum principal stress became along the radial direction. Therefore, the wound healing problem proves again that the cell orientation and alignment were determined by the local stress-state in the cell layer, demonstrating a typical stress-driven cell behavior.
4. Summary and outlook
In this post, we showed that the tension force universally presents and plays crucial roles in living organisms. And the living organisms have the ability to keep the tension at certain values for different structural levels, which is called tensional homeostasis, in contrast to chemical homeostasis. Tensional homeostasis should be an important mechanism of living organisms for optimizing their efficiency of functioning in different physiological activities. Deviation from the homeostasis will cause diseases. Therefore, it is possible to treat the diseases by tuning the tension back to their homeostasis values. The medicine based on this kind of concept should belong to mechanomedicine.
We showed that many cell behaviors depend on the tensional homeostasis, such as traction force distribution, cell polarization, and orientation. And these behaviors/properties influence the cell adhesion and migration, and then the morphogenesis of tissue and organs. It can be imagined that by tuning the magnitude and distribution of the tension in living organisms we can tune cell polarization, the cell-matrix interaction, and thus the tissue morphogenesis in the processes of treating the diseases.
In fact, there are many kinds of non-drug therapies, including those in Chinese Medicine, such as acupuncture, acupressure, and Tuina [28], which should be thought as typical forms of application of mechanomedicine. These therapies are effective, cheap and convenient. Currently, the underlying mechanisms of the efficacy of these precious wisdom presented by these treatments are largely unknown. Mechanomedicine may provide us an approach for disclosing these secrets.
References
[1] Bloomfield, A. L., "Some Problems of Peptic Ulcer," Am. J. Surg., vol. 89, pp. 1109-1115, 1955, doi: 10.1016/0002-9610(55)90472-4.
[2] Harsh, Mohan, "Textbook of Pathology." Jaypee Brothers Medical Publishers, Philadelphia 2014.
[3] Han, Y., Pegoraro, A. F., Li, H., Li, K., Yuan, Y., Xu, G., Gu, Z., Sun, J., Hao, Y., Gupta, S. K., Li, Y., Tang, W., Kang, H., Teng, L., Fredberg, J. J., Guo, M., "Cell swelling, softening and invasion in a three-dimensional breast cancer model," Nat Phys, vol. 16, pp. 101-108, 2020, doi: 10.1038/s41567-019-0680-8.
[4] Chen, B., Ji, B., Gao, H., "Modeling Active Mechanosensing in Cell-Matrix Interactions," Annual Review of Biophysics, vol. 44, pp. 1-32, 2015, doi: 10.1146/annurev-biophys-051013-023102.
[5] Du, S., Lutkenhaus, J., "At the Heart of Bacterial Cytokinesis: The Z Ring," Trends Microbiol., vol. 27, pp. 781-791, 2019, doi: 10.1016/j.tim.2019.04.011.
[6] Li, J., Cheng, L., Jiang, H., "Cell shape and intercellular adhesion regulate mitotic spindle orientation," Mol. Biol. Cell, vol. 30, pp. 2458-2468, 2019, doi: 10.1091/mbc.E19-04-0227.
[7] Shvartsman, Stanislav Y., Krajnc, Matej, "Nuclear (Bio) physics in the Embryo," Cell, vol. 177, pp. 799-801, 2019, doi: 10.1016/j.cell.2019.04.031.
[8] Lin, S., Li, B., Lan, G., Feng, X., "Activation and synchronization of the oscillatory morphodynamics in multicellular monolayer," Proc. Natl. Acad. Sci. U.S.A., vol. 114, pp. 8157-8162, 2017, doi: 10.1073/pnas.1705492114.
[9] Y. C, Fung, "Biomechanics: Motion, Flow, Stress and Growth.." Springer-Verlag, New York, 1990.
[10] Wang, N., Suo, Z., "Long-distance propagation of forces in a cell," Biochem. Biophys. Res. Commun., vol. 328, pp. 1133-1138, 2005, doi: 10.1016/j.bbrc.2005.01.070.
[11] Wei, F., Xu, X., Zhang, C., Liao, Y., Ji, B., Wang, N., "Stress fiber anisotropy contributes to force-mode dependent chromatin stretching and gene upregulation in living cells," Nat. Commun., vol. 11, p. 12, 2020, doi: 10.1038/s41467-020-18584-5.
[12] Siechen, S., Yang, S., Chiba, A., Saif, T., "Mechanical tension contributes to clustering of neurotransmitter vesicles at presynaptic terminals," Proc. Natl. Acad. Sci. U.S.A., vol. 106, pp. 12611-12616, 2009, doi: 10.1073/pnas.0901867106.
[13] Bu, B., Tian, Z., Li, D., Ji, B., "High Transmembrane Voltage Raised by Close Contact Initiates Fusion Pore," Front. Mol. Neurosci., vol. 9, p. 10, 2016, doi: 10.3389/fnmol.2016.00136.
[14] Park, J. S., Burckhardt, C. J., Lazcano, R., Solis, L. M., Isogai, T., Li, L. Q., Chen, C. S., Gao, B. N., Minna, J. D., Bachoo, R., DeBerardinis, R. J., Danuser, G., "Mechanical regulation of glycolysis via cytoskeleton architecture," Nature, vol. 578, pp. 621-626, 2020, doi: 10.1038/s41586-020-1998-1.
[15] Deguchi, S., Sato, M., "Biomechanical properties of actin stress fibers of non-motile cells," Biorh, vol. 46, pp. 93-105, 2009, doi: 10.3233/bir-2009-0528.
[16] Bershadsky, A. D., Balaban, N. Q., Geiger, B., "Adhesion-dependent cell mechanosensitivity," Annu. Rev. Cell. Dev. Biol., vol. 19, pp. 677-695, 2003, doi: 10.1146/annurev.cellbio.19.111301.153011.
[17] Kong, D., Ji, B., Dai, L., "Stabilizing to disruptive transition of focal adhesion response to mechanical forces," J. Biomech., vol. 43, pp. 2524-2529, 2010, doi: 10.1016/j.jbiomech.2010.05.019.
[18] Lu, L., Feng, Y., Hucker, W. J., Oswald, S. J., Longmore, G. D., Yin, F. C. P., "Actin stress fiber pre-extension in human aortic endothelial cells," Cell Motil. Cytoskeleton, vol. 65, pp. 281-294, 2008, doi: 10.1002/cm.20260.
[19] Arampatzis, A., Karamanidis, K., Albracht, K., "Adaptational responses of the human Achilles tendon by modulation of the applied cyclic strain magnitude," J. Exp. Biol., vol. 210, pp. 2743-2753, 2007, doi: 10.1242/jeb.003814.
[20] Bayraktar, H. H., Keaveny, T. M., "Mechanisms of uniformity of yield strains for trabecular bone," J. Biomech., vol. 37, pp. 1671-1678, 2004, doi: 10.1016/j.jbiomech.2004.02.045.
[21] Guo, X., Lu, X., Kassab, G. S., "Transmural strain distribution in the blood vessel wall," Am. J. Physiol. Heart Circ. Physiol., vol. 288, pp. H881-H886, 2005, doi: 10.1152/ajpheart.00607.2004.
[22] Harris, A. K., Wild, P., Stopak, D., "Silicone-rubber substrata - new wrinkle in the study of cell locomotion," Science, vol. 208, pp. 177-179, 1980, doi: 10.1126/science.6987736.
[23] Zhong, Y., Ji, B., "Impact of cell shape on cell migration behavior on elastic substrate," Biofabrication, vol. 5, p. 10, 2013, doi: 10.1088/1758-5082/5/1/015011.
[24] He, S., Su, Y., Ji, B., Gao, H., "Some basic questions on mechanosensing in cell-substrate interaction," J. Mech. Phys. Solids, vol. 70, pp. 116-135, 2014, doi: 10.1016/j.jmps.2014.05.016.
[25] He, S., Green, Y., Saeidi, N., Li, X., Fredberg, J. J., Ji, B., Pismen, L. M., "A theoretical model of collective cell polarization and alignment," J. Mech. Phys. Solids, vol. 137, p. 15, 2020, doi: 10.1016/j.jmps.2018.103860.
[26] Zhong, Y., Ji, B., "How do cells produce and regulate the driving force in the process of migration?," Eur. Phys. J. Spec. Top., vol. 223, pp. 1373-1390, 2014, doi: 10.1140/epjst/e2014-02196-8.
[27] He, S., Liu, C., Li, X., Ma, S., Huo, B., Ji, B., "Dissecting Collective Cell Behavior in Polarization and Alignment on Micropatterned Substrates," Biophys. J., vol. 109, pp. 489-500, 2015, doi: 10.1016/j.bpj.2015.06.058.
[28] Kaptchuk, T.J., "The Web That Has No Weaver." Congdon and Weed, Chicago, 1983.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}